![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
504 Cards in this Set
- Front
- Back
|
PORQUE É QUE AS CÉLULAS SÃO A UNIDADE BASE DA VIDA? |
Porque os seus compostos se degradam quando sozinhos, e quando são removidas(células) têm a capacidade de crescer e se reproduzir. |
|
|
CARACTERÍSTICAS DE TODAS AS CÉLULAS EUCARIOTICAS |
Complexas e organizadas; DNA; capazes de se multiplicar; consomem e produzem energia; armazenam enrgia como ATP; Realizam reações químicas; possuem um citosqueleto e atividade mecânica; respondem a estímulos; auto-regulam-se(morte celular no caso de não funcionamento. |
|
|
CARACTERÍSTICAS MOLECULARES DA ÁGUA |
Molécula polar(os eletroes são mais atraídos pelo oxigénio). No estado sólido há um decréscimo de densidade( devido as interaçoes entre moléculas). Devido a isto a água possui tensão superficial, permitindo a deslocação de certos animais. Dissolve moléculas polares. |
|
|
TIPOS DE MONOSSACÁRIDOS |
Frutose, glucose(alfa(carbono 1 abaixo do plano da molécula))(beta(carbono 1 acima do plano da molécula) |
|
|
TIPOS DE DISSACÁRIDOS |
Sacarose( Glucose + frutose ) Lactose ( Galactose + glucose ) |
|
|
POLISSACÁRIDOS |
Amido(é digerido)( Glucose alfa(amilose) + glucose alfa(amilopectina); Glicogénio(também glucose alfa); Celulose(não é digerido)(glucose beta) |
|
|
CARACTERÍSTICAS COMUNS DOS LÍPIDOS |
Normalmente hidrófobas, constituidas por ácidos gordos. Saturado- Sem ligaões duplas H-O; Insaturado: Ligaçoes duplas. |
|
|
TRIGLICÉRIDOS |
Ésteres de glicerol( 3 ácidos gordos) |
|
|
FOSFOLÍPIDOS |
Moléculas anfipáticas. 2 Cadeias de ácidos gordos+glicerol+grupo fosfato. Molécula polar. |
|
|
ESTERÓIDES |
Molécula hidrofóbica ou anfipática(colesterol) |
|
|
PROTEÍNAS(GERAL) |
São constituidas por três ou mais estruturas, só estando funcionais quando atigem a terceira estrutura. |
|
|
ESTRUTURA PRIMÁRIA(PROTEÍNAS) |
Aminoácidos |
|
|
ESTRUTURA SECUNDÁRIA (PROTEÍNAS) |
Existem dois tipos: Hélice alfa - Existem ligações de hidrogénio entre aminoácidos da mesma cadeia. Estrutura planar Beta - Ligaçoes entre aminoácidos de cadeias diferentes. |
|
|
ESTRUTURA TERCIÁRIA (PROTEÍNAS) |
Supra-enrolamento, há ligaçoes entre as várias colunas. A proteína fica funcional |
|
|
ESTRUTURA QUATERNÁRIA (PROTEÍNAS) |
Conjugação de unidades de estrutura terciária. |
|
|
ÁCIDOS NUCLEICOS |
Macromoléculas cuja unidade estrutural é o nucleótido. |
|
|
NUCLEOPROTEÍNAS |
Estruturas resultantes da associação de ácidos nucleicos com proteínas. |
|
|
TIPOS DE RNA |
RNA mensageiro, RNA de transferência, RNA ribossómico |
|
|
TRANSCRIÇÃO |
Após a replicação, a enzima RNA polimerase sintetiza RNA pré-mensageiro. Este é processado, sendo removidos os intrões. Fica um mRNA funcional. |
|
|
TRADUÇÃO |
A sub-unidade menor do ribossoma liga-se ao mRNA. Quando surge o codão de iniciação(AUG) une-se o tRNA com o anticodão AUC (met)(P), de seguida une-se a sub-unidade maior do ribossoma e há a elongação. Depois o tRNA com o anticodão complementar ao codão seguinte(A) liga-se e continua o processo, saindo o tRNA anterior. O processo termina quando o ribossoma atinge codão stop. |
|
|
PROCESSAMENTO |
Existem proteínas (chaperonas) que ajudam as proteínas a adquirir a sua estrutura terciária ou a manter a sua estrutura para passarem através de membranas. As chaperoninas(maiores que as chaperonas), funcionam como um tunel pela qual as proteínas passam e são processadas. |
|
|
DIFERENÇAS ENTRE A CÉLULA PROCARIOTA E EUCARIOTA |
Núcleo - ausente na procariota ; Dimensão - menor a procariota; Citosqueleto - ausente na procariota(podem possuir estruturas semelhantes); Organitos - ausentes na procariota(aceita-se como organito tudo excepto ribossomas). DNA - mais pequeno nos procariotas. Cromossomas - simples e circular nos procariotas vs múltiplos e lineares nos Eucariotas |
|
|
O QUE SÃO MICROESFERAS? |
São macromoléculas orgânicas envolvidas por membranas.
|
|
|
CARACTERÍSTICAS DA PRIMEIRA CÉLULA |
A primeira célula passa a possuir RNA auto-replicante dentro de um meio intracelular, envolvido por uma camada fosfolípida que o separa do meio extracelular. |
|
|
QUAL FOI A SEQUÊNCIA DA EVOLUÇÃO METABÓLICA? |
Glicólise - Fotossíntese - Metabolismo oxidativo(respiração aeróbica) |
|
|
PORQUE PODE OCORRER O METABOLISMO OXIDATIVO? |
Pois com a evolução da fotossíntese estava disponível oxigénio na atmosfera. |
|
|
QUAL A IMPORTÂNCIA DO APARECIMENTO DE ORGANITOS NOS EUCARIONTES? |
Possiblita a eficiência funcional das células, mesmo com uma grande dimensão. |
|
|
QUAL A TEORIA ATUALMENTE ACEITE PARA EXPLICAR O APARECIMENTO DE ORGANITOS CELULARES? |
Teoria Endossimbionte |
|
|
O QUE DIZ A TEORIA ENDOSSIMBIONTE? |
Que as células eucariotas evoluiram apartir de uma simbiose entre células procariotas, sendo os organelos células procariotas. |
|
|
DÁ DOIS EXEMPLOS DE SIMBIOSE ENTRE CÉLULAS PRÓCARIOTAS QUE ORIGINARAM CÉLULAS EUCARIÓTAS |
Bactérias aeróbicas - mitocôndrias . Cianobactérias- cloroplastos. |
|
|
QUAIS SÃO OS DADOS QUE APOIAM A TEORIA ENDOSSIMBIÓTICA? |
Os clorosplastos e mitocôndrias apenas se formam quando estão presentes na célula de origem. Os clpts e mtcrds possuem o seu próprio genoma e meio de síntese proteica e assemelha-se ao procariota. Existem antibióticos que só afetam os organelos e bactérias. |
|
|
QUAIS SÃO OS DOIS DIFERENTES REINOS DE BACTÉRIAS? |
Archae e Eubacteria |
|
|
TRÊS TIPOS DE EUBACTÉRIAS MAIS IMPORTANTES |
Micoplasmas; Fotossintéticas, saprófitas, parasitas; Cianobactérias |
|
|
CARACTERIZA AS EUBACTÉRIAS MICOPLASMAS |
São as bactérias mais simples, não possuem parede celular e são as menores |
|
|
CARACTERIZA AS CIANOBACTÉRIAS |
Grande capacidade de adaptação, possuem uma cor azulada, usam apenas água, luz e azoto atmosférico. |
|
|
QUE TIPO DE RIBOSSOMA SE ENCONTRA NAS CÉLULAS PROCARIOTAS? |
Ribossoma 70s |
|
|
QUAL O SIGNIFICADO DE UM RIBOSSOMA SER 70S OU 80S? |
Significa que demora 70 ou 80 segundos a sedimentar em centrifugação. |
|
|
QUE TIPO DE RIBOSSOMA SE ENCONTRA NAS CÉLULAS EUCARIOTAS? |
80s |
|
|
QUE FUNÇÕES VITAIS TEM A MEMBRANA CELULAR NOS PROCARIOTAS? |
Na membrana celular estão várias proteínas importantes para a troca de nutrientes e de energia(ATPases). |
|
|
QUE DOIS TIPOS DE BACTÉRIAS SE CONSEGUE DIFERENCIAR PELA PAREDE CELULAR? |
Bactérias Gram+ e Gram- |
|
|
QUAIS SÃO AS CARACTERÍSTICAS DIFERENCIAIS DAS BACTÉRIAS GRAM+? |
Possuem apenas uma membrana celular rodeada por uma parede celular espessa de peptidoglicanos. Contém cadeias de ácidos lipoteicos e teicoicos que interagem com o meio extracelular. Fixa o corante. |
|
|
QUAIS SÃO AS CARACTERÍSTICAS DIFERENCIAIS DAS BACTÉRIAS GRAM-? |
Possui duas membranas: celular e externa. Entre as duas membranas possui uma fina camada de peptidoglicanos. Parede celular=peptidoglicanos +membrana externa. Contém lipopolissacáridos(antigénios O) - comunicam com o meio extracelular. As lipoproteínas ligam-se aos peptidoglicanos - ligando a camada externa da parede celular à interna. |
|
|
FUNÇÃO E CONSTITUIÇÃO DA CÁPSULA DA BACTÉRIA |
Envolve a parede celular, protegendo a bactéria e impedindo que seja reconhecida por outras células. |
|
|
FUNÇÃO E CONSTITUIÇÃO DA CAMADA S DA BACTÉRIA |
Camada rígida constituida por proteínas que envolve a parede celular. |
|
|
FUNÇÃO E CONSTITUIÇÃO DO FLAGELO DA BACTÉRIA |
Filamento constituido por proteínas. O seu movimento rotativo garante locomoção. |
|
|
FUNÇÃO E CONSTITUIÇÃO DAS PILI OU FÍMBRIAS DA BACTÉRIA |
Filamentos proteicos, curtos e sem movimento. Têm função de reconhecimento e adesão. Reprodução parasexuada(transferência parcial de material genético). |
|
|
PORQUE SÃO MAIS EFICIENTES AS CÉLULAS EUCARIOTAS? |
Pois possuem mais organitos. |
|
|
O QUE É NECESSÁRIO PARA AS CÉLULAS PODEREM COMUNICAR? |
É necessário haver continuidade. |
|
|
O QUE POSSIBLITOU A MULTICELULARIDADE? |
A formação de colónias e a subsequente diferenciação e especialização. |
|
|
TIPOS DE TECIDOS CELULARES |
Epitelilal, conjuntivo, nervoso , muscular |
|
|
DEFINIÇÃO E FUNÇÃO DE TECIDO EPITELIAL |
Revestimento e proteção. São as células que cobrem as superfícies corporais. Na pele as células mortas fazem parte do epitélio. As células possuem uma proximidade tal que impedem a passagem de qualquer substância, para passarem substâncias têm de ser transportadas por transportadores especiais. |
|
|
DEFINIÇÃO E FUNÇÃO DE TECIDO CONJUNTIVO |
Tecido que une os epitélios. Ao contrário do tecido epitelial, existem grandes espaços intracelulares. Grande volume de matriz celular. |
|
|
DEFINIÇÃO E FUNÇÃO DE TECIDO NERVOSO |
Epitélio altamente modificado, neurónios + células da glia(nutrição dos neurónios). São as células que necessitam de mais energia. |
|
|
TIPOS DE TECIDO MUSCULAR |
Liso - involuntário( contrações lentas - maior duração). Estriado - voluntário( contrações rápida - curta duração) |
|
|
ORGANIZAÇÃO DE TECIDOS DAS PLANTAS |
Epiderme, tecido vascular, tecido cortical |
|
|
DEFINIÇÃO DA EPIDERME DA PLANTA |
Células que revestem a planta. Possuí uma camada protetora(cutícula). |
|
|
DEFINIÇÃO DO TECIDO VASCULAR DA PLANTA |
Xilema e floema. Tecidos de transporte. |
|
|
QUAIS SÃO AS CÉLULAS MAIS EFICAZES NA RESPIRAÇÃO? PROCARIOTAS OU EUCARIOTAS? |
Eucariotas |
|
|
QUAIS SÃO OS DOIS TIPOS DE FOTOSSÍNTESE BACTERIANA? |
Oxigénica e não-oxigénica. |
|
|
QUAL ABSORVE LUZ NUM MAIOR COMPRIMENTO DE ONDA? BACTERIOCLOROFILA-A OU CLOROFILA? |
Bacterioclorofila-A |
|
|
QUAL A IMPORTÂNCIA DAS CIENOBACTÉRIAS PARA O SURGIMENTO DAS PLANTAS? |
As cianobactérias fixam o azoto atmosférico - transformando-o em azoto orgânico que pode ser utilizado pelas plantas. |
|
|
CONSTITUIÇÃO DAS CIENOBACTÉRIAS |
Heterocistos, células fotossintéticas, estruturas membranares. |
|
|
QUAL É A FUNÇÃO DOS HETEROCISTOS NAS CIENOBACTÉRIAS? |
Fixação do azoto |
|
|
QUAL É A CARACTERÍSTICA DAS CIENOBACTÉRIAS QUE NÃO É BACTERIANA? |
Estruturas membranares. |
|
|
O QUE OCORREU NO PLANETA QUE PERMITIU O SURGIMENTO DAS CÉLULAS EUCARIOTAS? |
Oxigénio na atmosfera. |
|
|
A PAREDE CELULAR É COMPOSTA POR UM POLÍMERO. QUE POLÍMERO É ESTE? |
Celulose |
|
|
NA PAREDE CELULAR AS CADEIAS DE CELULOSE ASSOCIAM-SE. O QUE FORMAM? |
Microfibrilhas |
|
|
QUE OUTROS DOIS POLISSACARÍDEOS ESTÃO ENTRE AS MICOFIBRILHAS DE CELULOSE? |
Hemicelulose e pectina |
|
|
O QUE SÃO AS HEMICELULOSES E QUAL A SUA FUNÇÃO NA PAREDE CELULAR? |
São polissacarídeos muito ramificados que estão ligados através de pontes de hidrogénio à superfície da microfibrilhas. A sua função é estabilizar as micofibrilhas de forma a formarem uma fibra compacta. |
|
|
O QUE SÃO AS PECTINAS E QUAL A SUA FUNÇÃO? |
São políssacarídeos ramificados que contém um grupo ácido. Permitem ligações com catiões, devido ao seu caracter ácido. Permitem também a absorção de água formando uma massa gelatinosa. |
|
|
O QUE EXISTE ENTRE AS CÉLULAS DAS PLANTAS ?QUAL A SUA FUNÇÃO? |
Lamela média, funciona como adesivo entre células e é formada por pectinas |
|
|
QUE ENZIMAS ATUAM NA EXPANSÃO DA PAREDE CELULAR? QUAL A SUA FUNÇÃO? |
As expansinas. Libertam as ligações entre micofibrilhas permitindo a expansão da parede celular. |
|
|
QUE HORMONAS ATIVAM AS EXPANSINAS? |
Auxinas |
|
|
O QUE PERMITE A LIBERTAÇÃO DA LIGAÇÕES ENTRE MICROFIBRILHAS PROMOVIDA PELAS EXPANSINAS? |
Permite que a célula expanda. A pressão de turgescência(pressão osmótica) faz entrar água no vacúlo que aumenta do tamanho e causa a expansão da célula e da parede celular. |
|
|
O QUE SE SUCEDE À PAREDE CELULAR DURANTE A SUA EXPANÇÃO? |
São depositados os seus constituientes à medida que se expande. |
|
|
QUE DIFERENÇAS POSSUI A PAREDE CELULAR SECUNDÁRIA FACE À PRIMÁRIA? |
Não possui pectina e possui 50-80% celulose, sendo mais rígida, também um polímero chamado de lignina que contribui para a sua rigidez. Na parede celular secundária as microfibrilhas estão muito organizadas, em contraste com a parede celular primária. |
|
|
QUANDO OCORRE A FORMAÇÃO DA PAREDE CELULAR SECUNDÁRIA? |
Quando já não há expansão da primária. |
|
|
O QUE DETERMINA A ORIENTAÇÃO DO ALONGAMENTO DA CÉLULA? |
A orientação das microfibrilhas |
|
|
DE QUE FORMA OCORREM TROCAS ENTRE CÉLULAS QUANDO A PAREDE CELULAR É ESPESSA? |
Através de pontuações, zonas onde durante a diferenciação tecidular não ocorreu deposição de novo material. |
|
|
O QUE É A LENHINA? |
É um polímero fenólico. |
|
|
QUAL A CONSEQUÊNCIA DA LENHIFICAÇÃO DA PAREDE CELULAR? |
Substituição de componentes da matriz pro lenhina, conferindo mais rigidez à parede celular. |
|
|
QUE FUNÇÕES PERMITE A LENHIFICAÇÃO DA PAREDE CELULAR? PORQUÊ? |
Funções de suporte e de condução(promovido pelo facto da lenhina ser hidrófoba). Permite o porte ereto das plantas e caracteriza as plantas vasculares. |
|
|
O QUE SE SUCEDE A UMA CÉLULA AO SER LENHIFICADA? O QUE PERDE? |
A célula morre, fica sem protoplasma |
|
|
ONDE SE PODE ENCONTRAR LENHICAÇÃO CELULAR? QUAL A SUA IMPORTÂNCIA? |
No xilema, impede que este perda água |
|
|
EM QUE CONSISTE A SUBERIFICAÇÃO? |
Deposição de camadas de suberina no interior da parede celular. |
|
|
QUE FUNÇÃO TEM A SUBERIFICAÇÃO? |
Proteje e impermiabiliza. |
|
|
O QUE SE SUCEDE A UMA CÉLULA APÓS A SUA SUBERIFICAÇÃO? |
Ocorre a morte desta célula. |
|
|
EM QUE CÉLULAS SE PODE ENCONTRAR SUBERIFICAÇÃO? |
Na rizoderme e nas células da endoderme. |
|
|
QUAL A COMPOSIÇÃO DA CUTÍCULA VEGETAL QUE PROTEGE AS PAREDES CELULARES? |
Cutina -polímero de ácidos gordos insoluvel e por ceras lípidicas. |
|
|
EM QUE CONSISTE A CUTINIZAÇÃO? |
Na deposição de ceras e cutina na parede celular externa. |
|
|
QUE VANTAGENS TEM A CUTINIZAÇÃO? |
Impermiabiliza. Protege as plantas contra radiação excessiva, regula o grau de hidratação, impede a invasão de agentes patogénicos. |
|
|
ONDE OCORRE A CUTINIZAÇÃO? |
Limita-se à parede celular das células da epiderme. |
|
|
DE QUE FORMA SE INICIA A FORMAÇÃO DA PAREDE CELULAR? |
Quando um sistema de microtubulos chamado de fragmoplasto orienta as vesículas do complexo de Golgi e do Retículo Endoplasmático a fundirem-se no interior da célula. |
|
|
COMO SE CHAMA A ESTRUTURA QUE SE FORMA NO INTERIOR DA CÉLULA QUE CONTÉM AS CÉLULAS PERCURSORAS DA FORMAÇÃO DA PAREDE CELULAR? |
Placa celular. |
|
|
COMO SE FORMA A PAREDE CELULAR A PARTIR DA PLACA CELULAR? |
No local onde está a placa celular há fusão de mais vesículas, contendo hemilocelulose e pectina orientada pelos microtubulos. Neste mesmo local ficará a lamela média. |
|
|
QUAL A ENZIMA QUE SINTETIZA A CELULOSE? |
Celulose sintase, ou roseta. |
|
|
O QUE ACONTECE APÓS A SÍNTESE DAS MICROFIBRILHAS DE CELULOSE PELA ENZIMA ROSETTA? |
A celulose é tranlocada para o exterior da célula(para a parede celular), através de um poro criado por sub-unidades da enzima rosetta. De seguida os microtubulos orientam as microfibrilhas definindo a elongação da célula. |
|
|
QUE TRÊS COMPONENTES COMPÕEM A MATRIZ EXTRACELULAR ANIMAL? |
Fibras proteicas; rede polissacarídia gelificada; proteínas de ligação |
|
|
QUAIS SÃO AS DUAS PRINCIPAIS PROTEÍNAS DA MATRIZ EXTRACELULAR ANIMAL QUE CONSTITUEM AS FIBRAS PROTEICAS? |
Colagénio e elastina. |
|
|
COMO É A ESTRUTURA DO COLAGÉNIO? |
O colagénio é composto por três cadeias polipetidicas entrelaças em hélice. |
|
|
QUE NOME TÊM AS CADEIAS POLIPEPTÍDICAS QUE CONSTITUEM A TRIPLA HÉLICE DE COLAGÉNIO? |
São cadeias alfa. |
|
|
COMO É CONSTITUIDA CADA CADEIA ALFA QUE COMPÕEM A TRIPLA HÉLICE DE COLAGÉNIO? |
Um aminoácido de glicina de três em três posições, prolina(X) e hidroxiprolina(Y). Sendo a estrutura normalmente: Gly-X-Y-Gly-X-Y(....) |
|
|
QUE FUNÇÃO TEM A GLICINA NA TRIPLA HÉLICE DE COLAGÉNIO? |
Sendo o aminoácido mais pequeno, composto em parte só po hidrogénio, tem uma função essencial na junção das cadeias alfa de modo a que estas estejam próximas o suficiente para formar a tripla hélice. |
|
|
PORQUE RAZÃO ESTABILIZAM A ESTRUTURA EM HÉLICE DO COLAGÉNIO A PROLINA E A HIDROXIPROLINA? |
Pois a sua estrutura molecular é um anel. |
|
|
ONDE SE INICIA A FORMAÇÃO DAS FIBRILHAS DE COLAGÉNIO? |
No lúmen do retículo endoplasmático. |
|
|
DURANTE A FORMAÇÃO DAS FIBRILHAS DE COLAGÉNIO, O QUE SE SUCEDE NO RETÍCULO ENDOPLASMÁTICO? |
Formam-se as cadeias percursoras de colagénio, a prolina é modificada para hidroxiprolina. De seguida forma-se o procolagénio no interior de vesículas, que contém partes que não estão em hélice. |
|
|
DURANTE A FORMAÇÃO DE FIBRAS DE COLAGÉNIO, O QUE SE SUCEDE APÓS A FORMAÇÃO DE VESÍCULAS NO RETÍCULO COM PROCOLAGÉNIO? |
Estas vesículas libertam-se e fundem-se com a membrana da célula, depois o procolagénio passa para o espaço extracelular e as peptidases clivam o procolagénio. |
|
|
APÓS A CLIVAGEM DE MOLÉCULAS DE PROCOLAGÉNIO E FORMAÇÃO DE COLAGÉNIO, COMO SE FORMAM AS FIBRAS? |
O colagénio une-se em fibrilhas que são fortacelidas por ligações covalentes entre as diferentes moléculas. Estas fibrilhas associam-se depois umas ás outras para formar fibras de colagénio que são depositadas no meio extracelular. |
|
|
O QUE SÃO OS FIBROBLASTOS? ONDE SÃO MAIS COMUNS? |
São células que sintetizam colagénio e mariz extracelular. São mais comuns no tecido conjuntivo. |
|
|
O QUE É A SÍNDROME DE EHLERS-DANLOS? |
É uma síndrome que resulta de uma mutação que causa um defeito na síntese de colagénio. Resulta em hipermobilidade articular. |
|
|
QUAL É A PROTEÍNA PRINCIPAL DAS FIBRAS DA MATRIZ EXTRACELULAR ELÁSTICA? |
Elastina |
|
|
COMO É CONSTITUIDA A ELASTINA? |
Por tropoelastina e lisil-oxidase. |
|
|
ONDE É MAIS COMUM ENCONTRAR ELASTINA? |
Em orgãos realizam contrações(p.e. pulmões). |
|
|
QUE FUNÇÃO TEM A ELASTINA? |
Funciona como um elástico contraindo e esticando consoante o necessário. |
|
|
QUE DOIS ELEMENTOS COMPÕEM A REDE POLISSACARÍDICA GELIFICADA? |
Glisoaminoglicanos(GAGs) e proteoglicanos. |
|
|
COMO SÃO COMPOSTOS OS GLISOAMINOCLICANOS? |
Repetição de unidades de dissacarídeos. |
|
|
DIZ O NOME DE UM GAG |
Ácido hialuranoico |
|
|
QUE FUNÇÃO TÊM OS GAGs? |
Dão suporte mecânico à matriz extracelular. Sendo carregados negativamente, ligam-se a catiões e aprisionam moléculas de água formando um gel. |
|
|
QUAL É O ÚNICO GAG QUE EXISTE NA FORMA DE UMA ÚNICA CADEIA DE POLISSACARÍDEOS? AO QUE SE LIGA? |
É o ácido hialuranoico e liga-se a proteínas formando proteoglicanos. |
|
|
QUAL É O PROTEOGLICANO MAIS COMUM NA CARTILAGEM? |
O agrecano |
|
|
QUAIS SÃO AS PROTEÍNAS DE LIGAÇÃO MAIS COMUNS NA MATRIZ EXTRACELULAR ANIMAL? |
Fibronectinas, lamininas e entactinas. |
|
|
QUAL É A PRINCIPAL PROTEÍNA DE LIGAÇÃO DO TECIDO CONJUNTIVO? |
Fibronectina. |
|
|
COMO É COMPOSTA A FIBRONECTINA? |
É uma glicoproteína composta por duas cadeias de polipeptidos. |
|
|
A QUE ESTRUTURAS SE LIGA A FIBRONECTINA? |
Liga-se a fibrilhas de colagénio e GAGs, ligando também estes uns aos outros. |
|
|
A FIBRONECTINA POSSUI UM LOCAL COM FUNÇÕES IMPORTANTES A NÍVEL CELULAR, QUAL É ? |
Possui um local que é responsavel pela ligação a recetores da superficie da célula ás fibronectinas. Assim permite a ligação entre componentes da matriz extracelular e a superfície das células. |
|
|
A LÂMINA BASAL CONTÉM PROTEÍNAS DE LIGAÇÃO ESPECÍFICAS. QUAIS SÃO? |
São as lamininas
|
|
|
COMO SÃO CONSTITUÍDAS AS LAMININAS? |
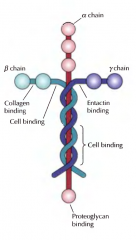
Compostas por três cadeias: alfa, beta e gama. Têm locais de ligação a proteínas receptoras(como as integrinas) e estão associadas a outras proteínas de ligação, as entactinas. |
|
|
QUAIS SÃO OS RECETORES MEMBRANARES MAIS IMPORTANTES NA LIGAÇÃO DA MEMBRANA CELULAR À MATRIZ EXTRACELULAR? |
São as integrinas |
|
|
COMO SÃO COMPOSTAS AS INTEGRINAS? |
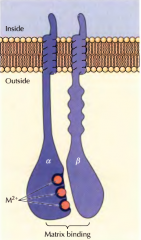
Por duas sub-unidades: alfa e beta. |
|
|
QUAL É UMA DAS VANTAGENS DA FLUIDEZ MEMBRANAR? |
Permite interações de curta duração |
|
|
QUAIS SÃO OS PRINCIPAIS CONSTITUITES DA MEMBRANA CELULAR EUCARIÓTICA? |
Lípidos, proteínas e hidratos de carbono. |
|
|
QUAIS SÃO OS TRÊS PRINCIPAIS LÍPIDOS DA MEMBRANA CELULAR? |
Fosfoglicéridos, esfingolípidos e esterois |
|
|
DÁ UM EXEMPLO DE UM FOSFOGLICÉRIDO, DE UM ESFINGOLÍPIDO E DE ESTEROL |
Fosfatidilcolina, esfingomielina e o colesterol. |
|
|
O QUE É COMUM ENTRE TODAS OS LÍPIDOS MEMBRANARES EUCARIOTAS? |
São moléculas anfipáticas( uma região hidrofílica e outra hidrofóbica). |
|
|
QUAIS SÃO OS QUATRO FOSFOLÍPIDOS MAIS IMPORTANTES DA MENBRANA CELULAR EUCARIÓTICA? |
Fosfatidilcolina, fosfatidiletanolamina, fosfatidilserina e esfingomielina. |
|
|
COMO ESTÃO DISTRIBUIDOS OS DIFERENTES FOSFOLÍPIDOS PELA MEMBRANA CELULAR EUCARIOTA? |
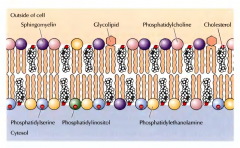
Assimétricamente. Na parte exterior estão a fosfatidilcolina e a esfingomielina, na parte interior estão a fosfatidiletanolamina e a fosfatidilserina. |
|
|
O QUE GARANTE A ASSIMETRIA DA MEMBRANA PLASMÁTICA EUCARIOTA? |
Garante a funcionalidade através da presença de cargas diferentes. |
|
|
O QUE DETERMINA A FLUIDEZ DA MENBRANA CELULAR EUCARIOTICA? |
O grau de insaturação dos ácidos gordos da extremidade hidrofóbica dos fosfolípidos, esta insaturaturação(ligações duplas) faz com que a membrana esteja num estado de gel semi-cristalino que lhe dá fluidez. |
|
|
COMO É A COMPOSIÇÃO GERAL DE UM FOSFOLÍPIDO? |
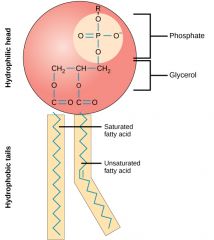
Cabeça hidrofílica(fosfato + glicerol) ; Extremidade hidrofóbica( ácidos gordos saturados e insaturados) |
|
|
O QUE PERMITE AOS FOSFOLÍPIDOS A FLUIDEZ MEMBRANAR? |
Flexão, movimentos laterais(difusão lateral), movimentos de abano, rotações e flip-flop. |
|
|
QUE FATORES ALTERAM A FLUIDEZ MEMBRANAR? |
A temperatura; a presença de colesterol, número de cadeias de ácidos gordos insaturadas. |
|
|
COMO VARIA A FLUIDEZ MEMBRANAR COM A TEMPERATURA? |
Se a temperatura for baixa o número de cadeias de ácidos gordos insaturadas ajuda a manter a fluidez, e o colesterol impede a compactação dos lípidos. A temperaturas altas o colesterol torna a membrana menso fluida |
|
|
COMO ESTÁ SITUADO O COLESTEROL NA BICAMADA FOSFOLÍPIDICA? |
Está entre os fosfolípidos com o seu grupo hidroxilo junto á cabeça hidrofílica dos fosfolípidos. |
|
|
O QUE FORMAM OS ESFINGOLÍPIDOS E O COLESTEROL NA MEMBRANA PLASMÁTICA? |
Jangadas lípidicas que se movem lateralmente entre os fosfolípidos associando-se a proteínas específicas. |
|
|
QUAIS SÃO OS TRÊS TIPOS PRINCIPAIS DE PROTEINAS DA MEMBRANA PLASMÁTICA EUCARIOTA? |
Proteínas integrais, periféricas e ancoradas. |
|
|
ONDE SE SITUAM AS PROTEÍNAS INTEGRAIS, PERIFÉRICAS E ANCORADAS? |
As integrais são transmembranares(situam-se no interior da membrana, entre os fosfolípidos), as periféricas estão totalmente fora da membrana estando apenas associadas a proteínas integrais. As proteínas ancoradas estão ligadas covalentemente a lípidos, diretamente ou indiretamente através do GPI(glicosilfosfatidilinositol) |
|
|
O QUE É O GPI(glicosilfosfatidilinositol)? |
São glicolípidos que se ligam covalentemente a proteínas e a ancoram á membrana plasmática. |
|
|
DENTRO DAS PROTEÍNAS TRANSMEMBRANARES EXISTEM DOIS TIPOS, QUAIS SÃO? |
Unipasse e multipasse. |
|
|
DESTINGUE AS PROTEÍNAS MULTIPASSE DAS PROTEÍNAS UNIPASSE. |
As proteínas unipasse apenas têm um segmento transmenbranar, enquanto as proteínas multipasse têm vários. |
|
|
QUE ESTRUTURA ADQUIREM NORMALMENTE AS PROTEÍNAS TRANSMEMBRANARES? |
Estruturas em hélice - alfa. Possuem zonas hidrofóbicas que atravessam na totalidade a bicamada fosfolípidica |
|
|
COMO OCORRE A LIBERTAÇÃO DE UMA PROTEÍNA INTEGRAL DA BICAMADA FOSFOLÍPIDICA? |
É necessário haver ruptura da bicamada, estas proteínas ficam associadas a lípidos. |
|
|
COMO OCORRE A LIBERTAÇÃO DE UMA PROTEÍNA PERIFÉRICA OU ANCORADA DA BICAMADA FOSFOLÍPIDICA? |
Através de processos que não alteram a membrana ficando estas proteínas livres, não presas a lípidos. |
|
|
COMO É A SOLUBILIDADE DA PROTEÍNAS DA MEMBRANA PLASMÁTICA? |
São soluveis em água. |
|
|
COMO É A ESTRUTURA DAS PORINAS? |
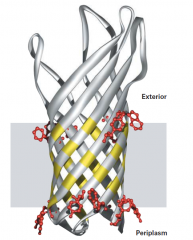
As porinas são proteínas integrais viradas do avesso. São constituidas por filamentos beta que formam uma estrutura semelhante a um tubo. Esta estrutura é hidrofóbica no exterior e hidrofílica no interior. Permitem assim a passagem de água. |
|
|
ONDE SE SITUAM PREFERENCIALMENTE AS PROTEÍNAS ANCORADAS AOS GPI? |
Nas jangadas lípidicas. |
|
|
ONDE SE SITUAM SEMPRE OS HIDRATOS DE CARBONO MEMBRANARES? |
No lado externo da membrana plasmática, associados a proteínas- glicoproteínas; ou a lípidos - glicolípidos. |
|
|
COMO SÃO GERALMENTE OS HIDRATOS DE CARBONO MEMBRANARES? |
São oligossacáridos ramificados com 15 unidades de monómeros. |
|
|
O QUE É O GLICOCÁLICE? |
É uma cobertura carbohidratada formada por oligossacarídeos de glicolípidos e por glicoproteínas transmembranares. |
|
|
COMO SE FORMA O GLICOCÁLICE? |
Forma-se quando hidratos de carbono membranares absorvem água.(hidratam-se) |
|
|
DIZ UM LOCAL ONDE SE PODE ENCONTRAR GLICOCÁLICE? |
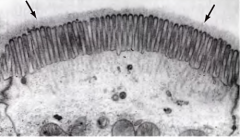
Por exemplo nas células epiteliais, no exterior das microvilosidades. |
|
|
QUAL É A FUNÇÃO DO GLICOCÁLICE? |
Protege a superfície celular. Os glícidos constituintes desempenham funções de marcadores celulares |
|
|
COMO É QUE OS HIDRATOS DE CARBONO MEMBRANARES DETERMINAM O GRUPO SANGUÍNEO? |
Existem três tipos de antigénios A, B, e AB que são oligossacarídeos(glicolípidos)que se ligam á membrana dos glóbulos vermelhos. Uma pessoa com antigénios B produz anticorpos anti-A. Uma pessoa sem antigénios é do grupo 0 e produz anticorpos B e A. |
|
|
COMO É QUE OS ANTIGÉNIOS DO GRUPO SANGUÍNEO DIFEREM UNS DOS OUTROS? |
Na porção terminal destes varia de glicolípido para outro na sequência de aminoácidos. |
|
|
ONDE OCORRE TRANSPORTE TRANSMEMBRANAR? |
Na membrana plasmática e nas membranas de organitos no interior das células. |
|
|
PORQUE RAZÃO A COMPOSIÇÃO INTERNA DA CÉLULA É MANTIDA SEMPRE IGUAL? |
Pois a membrana plasmática é seletivamente permeavel a pequenas moléculas. |
|
|
QUE SUBSTÂNCIAS CONSEGUEM ATRAVESSAR A BICAMADA FOSFOLÍPIDICA E QUAIS NÃO CONSEGUEM? |
Os gases, moléculas hidrofóbicas como o benzeno e pequenas moléculas polares como a água e o etanol conseguem. Moléculas plares grandes como a glucose e moléculas com carga como iões e amino acidos não conseguem. |
|
|
O QUE DETERMINA QUE UM TRANSPORTE TRANSMEMBRANAR SEJA PASSIVO? |
Ser a favor do gradiente de concentração: do mais concentrado para o menos; ou ser a favor do potencial eletroquímico: de onde há mais eletrões para onde há menos. Não haver consumo de energia |
|
|
O QUE É A DIFUSÃO PASSIVA? |
É o tipo de transporte membranar através do qual as substâncias como gases, moléculas hidrofóbicas e pequenas moléculas polares se difundem na bicamada fosfolípidica, segundo as regras do transporte passivo. |
|
|
QUE NOME TEM A DIFUSÃO PASSIVA DA ÁGUA? |
Osmose |
|
|
QUE NOME TEM UMA SOLUÇÃO EM QUE SE ESTIVER UMA CÉLULA OCORRE OSMOSE? E QUE NÃO OCORRE? |
Hipotónica(ganha água a célula); hipertónica(a célula perde água). Isotónica, nada acontece. |
|
|
QUAL É A DIFERENÇA ENTRE A DIFUSÃO FACILITADA E A DIFUSÃO PASSIVA? |
Na difusão facilitada as moléculas passam através de proteínas e na difusão simples através da bicamada fosfolípidica. |
|
|
QUE MOLÉCULAS É POSSIVEL TRANSPORTAR PARA O INTERIOR DA CÉLULA ATRAVÉS DA DIFUSÃO FACILITADA QUE NÃO ERA POSSÍVEL ATRAVÉS DA DIFUSÃO PASSIVA? |
Moléculas polares e com carga. |
|
|
QUAIS SÃO OS DOIS TIPOS DE PROTEÍNAS QUE DESEMPENHAM A DIFUSÃO FACILITADA? |
Proteínas transportadoras e proteínas canal. |
|
|
COMO REGULAM A ENTRADA DE MOLÉCULAS AS PROTEÍNAS TRANSPORTADORAS? |
Ligam-se a moléculas específicas e alteram a sua forma permitindo a passagem da molécula. |
|
|
COMO REGULAM A ENTRADA DE MOLÉCULAS AS PROTEÍNAS CANAL? |
Formam poros na membrana autorizando a difusão de moléculas de características e tamanhos específicos. |
|
|
DE QUE MANEIRA É A GLUCOSE TRANSPORTADA PARA O INTERIOR DA CÉLULA ATRAVÉS DA MEMBRANA? |
Por proteínas transportadoras. |
|
|
DÁ UM EXEMPLO DE UMA PROTEÍNA TRANSPORTADORA |
As permeases |
|
|
DÁ UM EXEMPLO DE UMA PROTEÍNA CANAL
|
As aquaporinas |
|
|
DIZ UM TIPO DE TRANSPORTE PASSIVO QUE ESTÁ DEPENDENTE DE PROTEÍNAS CANAL |
Transporte de iões |
|
|
QUE TIPOS DE CANAIS DE IÕES CONHECES QUE SÃO TRANSPORTE PASSIVO? |
Canais de iões que dependem de ligandos e dependentes de voltagem. |
|
|
COMO FUNCIONAM OS CANAIS DE IÕES DEPENDENTES DE LIGANDOS(TRASPORTE PASSIVO)? |
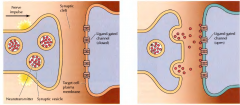
Uma molécula(ligando) liga-se a um local na proteína que faz abrir o canal permitindo a passagem de iões potássio e sódio. |
|
|
COMO FUNCIONAM OS CANAIS DE IÕES DEPENDENTES DE VOLTAGEM(TRASPORTE PASSIVO)? |
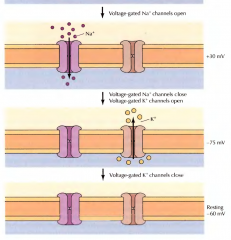
Uma mudança de potencial desecadeia a abertura do canais de sódio, entrando sódio no interior da membrana. De seguida a bomba de sódio fica inativa e dá-se a saída de potássio para o exterior da membrana para repor os valores de repouso. |
|
|
DURANTE A TRANSMISSÃO DE UM IMPULSO NERVOSO COMO ESTÃO CONCENTRAÇÕES DOS IÕES NUM POTENCIAL REPOUSO, DENTRO E FORA DA MEMBRANA DO AXÓNIO? |
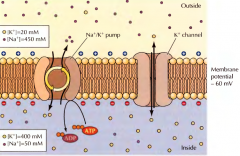
No interior há maior concentração de potássio e no exterior de sódio. O sódio é bombeado para fora enquanto o potássio para dentro. Durante este potencial a membrana é mais permeavel ao potássio. |
|
|
NO ESTADO DE REPOUSO, COMO SÃO AS CARGAS FORA E DENTRO DA MEMBRANA? |
Dentro são negativas e fora são positivas. |
|
|
DE QUE FORMA SE DÁ O TRANSPORTE ATIVO? |
Contra o gradiente de concentração ou potencial eletroquímico e com gasto de energia. |
|
|
DE QUE FORMA É OBTIDA ENERGIA NO TRANPORTE ATIVO DIRETO PRIMÁRIO? |
Utilizando o ATP de forma direta. (hidrósile) |
|
|
DÁ UM EXEMPLO DE TRANSPORTE ATIVO DIRETO PRIMÁRIO |
As bombas de potássio e sódio. |
|
|
QUE IMPORTÂNCIA TEM O TRANSPORTE ATIVO? |
Permite manter difereças de concentração crucias para a sobrevivência das células. |
|
|
EXPLICA COMO FUNCIONA A BOMBA DE SÓDIO-POTÁSSIO |
Primeiro( no interior da célula) o sódio liga-se á proteína(bomba) em locais esfecíficos, que estimula a hidrólise do ATP em ADP e a fosforilização da bomba. Esta fosforilização resulta em alterações na estrutura da proteína(bomba) que resultam na libertação do sódio para o meio extracelular. Ao mesmo tempo o potássio liga-se á bomba e dá-se a hidrólise do grupo fosfato(do ATP) que ficou fixo á bomba, o potássio é libertado para o meio intracelular. |
|
|
QUANTOS POTÁSSIOS E SÓDIOS SÃO TRANSPORTADOS EM CADA CICLO DA BOMBA DE SÓDIO-POTÁSSIO? QUANTAS MOLÉCULAS DE ATP SÃO USADAS? |
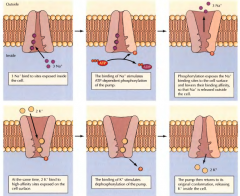
Três sódios e dois potássios, com uso de uma molécula de ATP |
|
|
QUE IMPORTÂNCIA TEM A BOMBA DE SÓDIO-POTÁSSIO? |
Tem importância na propagação de sinais nos músculos e nos nervos e no equílibrio omiostático e volume celular. |
|
|
PORQUE OUTRO NOME SÃO CONHECIDAS AS BOMBAS DE SÓDIO/POTÁSSIO CÁLCIO(ETC)? |
ATPases. |
|
|
QUAL É A MAIOR FAMILIA DE TRANPORTADORES MEMBRANARES? DE ONDE DERIVAM O SEU NOME? |
São as bombas tipo ABC. Derivam o seu nome do dominio extra-membranar de ligação ao ATP: ATP-binding cassete. |
|
|
COMO FUNCIONAM AS BOMBAS TIPO ABC? |
Uma proteína liga-se á molécula que vai ser transportada e leva-a até á bomba. As interações entre esta proteína e a bomba levam a alterações na bomba e nos seus dominios extra-membranares de ligação ao ATP. Desta forma a bomba hidrolisa duas moléculas de ATP, abrindo a bomba e permitindo a passagem da molécula. |
|
|
QUE TIPO DE SUBSTÂNCIAS TRANSPORTAM AS BOMBAS TIPO ABC? |
Vários tipos de nutrientes e biomoléculas, iões como (Cl-) e também transportam substâncias tóxicas para o exterior da célula. |
|
|
COMO FUNCIONAM AS BOMBAS TIPO V E F( O QUE TÊM EM COMUM)? |
São bombas constituidas por muitas sub-unidades. Em nenhuma delas a proteína transportadora(bomba) fica fosforilizada, transportam ambas apenas protões. |
|
|
COMO FUNCIONAM AS BOMBAS TIPO V? ONDE EXISTEM? |
As bombas tipo V hidrolizam o ATP para transportar protões contra o gradiente de cocentração. Estão presentes por exemplo nos lisossomas. |
|
|
COMO FUNCIONAM AS BOMBAS TIPO F? ONDE EXISTEM? |
Usam a energia dos protões que transportam para sintetizar ATP a partir de ADP. Existem por exemplo na membrana interna da mitocôndria. |
|
|
COMO SE DÁ O TRANPORTE ATIVO INDIRETO( COTRANSPORTE)? |
Utilizando a energia gerada pelo fluxo de iões a favor do gradiente. |
|
|
QUE TIPOS DE BOMBAS DE COTRANSPORTE EXISTEM? |
Bombas simporte e bombas antiporte. |
|
|
DÁ UM EXEMPLO DE UMA BOMBA SIMPORTE DE DO SEU FUNCIONAMENTO |
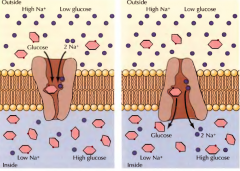
A bomba de sódio/Glucose. O transporte de iões sódio a favor do gradiente de concentração porporciona a energia para o transporte de glucose contra o gradiente permitindo a acumulação de glucose. |
|
|
DÁ UM EXEMPLO DE UMA BOMBA ANTIPORTE E DO SEU FUNCIONAMENTO
|
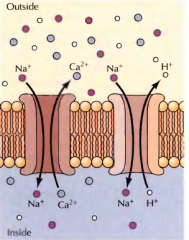
A bomba de sódio/cálcio. O cálcio e o hidrogénio são exportados das células o que porporciona energia para transportar sódio para o interior. |
|
|
EM QUE CONSISTE A FIBROSE CISTICA( A NIVEL CELULAR E DE SINTOMAS)? |
Resulta de uma mutação que tem como consequência a mal formação de bombas do tipo ABC e um transporte defeitouso de cloro (Cl-) para o exterior da células epiteliais. Resultam na obstrução das vias respiratórias com muco. |
|
|
QUE APARELHOS SÃO UTILIZADOS DURANTE A BIOGÉNESE MEMBRANAR? |
O complexo de golgi, o retículo endoplasmático e vesículas. |
|
|
QUAL É A FUNÇÃO DO RETÍCULO ENDOPLASMÁTICO RUGOSO? |
Sintetiza proteínas e armazena-as em vesículas. Vesículas que habitualmente se dirigem para o complexo de golgi. |
|
|
QUAL É A FUNÇÃO DO RETÍCULO ENDOPLASMÁTICO LISO? |
Sintetiza lipidos e armazena-os em vesículas de transporte. |
|
|
QUAL É A FUNÇÃO DAS VESÍCULAS DE TRANSPORTE? |
Transportam proteínas e lípidos para vários locais, entre os quais o complexo de Golgi. |
|
|
QUAL É A FUNÇÃO DO COMPLEXO DE GOLGI? |
Modifica lípidos e proteínas provenientes do retículo endoplasmático. Armazena-as em vesículas. |
|
|
QUAL É A FUNÇÃO DAS VESÍCULAS SECRETORAS? |
Transportam o que resulta do complexo de golgi e fundem-se com a membrana plasmática, ocorrendo as secreção. |
|
|
QUAL É A FUNÇÃO DOS LISOSSOMAS? |
Contéem enzimas digestivas que desintegram substâncias no interior da membrana. Por vezes fundem-se com outras vesículas e digerem o que está no seu interior. |
|
|
O QUE É O RETÍCULO ENDOPLASMÁTICO? |
É o maior organelo das células eucrioticas. Consiste numa rede de túbulos e cisternas que se extende a partir da membrana nuclear prelongando-se pelo citolplasma. Está coberto por uma membrana. |
|
|
COMO É CONSTITUIDO O RETÍCULO ENDOPLASMÁTICO? |
Pelo retículo endoplasmático rugoso(coberto de ribossomas), retículo de transição(de onde as vesículas saem), retículo liso(envolvido no metabolismo de lípidos. |
|
|
QUE DUAS PROPRIEDADES TÊM O RETÍCULO LISO E O RUGOSO? |
São maleaveis, podem alternar entre um e outro. |
|
|
O QUE É O COMPLEXO DE GOLGI? |
É o local onde as proteínas que vêm do retículo endoplasmático são processadas e divididas por endossomas, lisossomas, inseridas na membrana plasmática ou secretadas. Também é o local onde são sintetizados glicolipidos e esfingolipidos; e no caso das plantas, polissacarídeos da parede celular. |
|
|
COMO É DIVIDO O COMPLEXO DE GOLGI? |
É composto por cisternas e vesículas associadas. Possiu polaridade: uma face cis junto ao retículo e uma fase trans, de onde saem as proteínas. |
|
|
QUAL É O SENTIDO DE SECREÇÃO DAS PROTEÍNAS? |
RER->Golgi->vesículas secretoras -> exterior celular. |
|
|
DE QUE DUAS FORMAS PODEM AS PROTEÍNAS SER TRANSLOCADAS PARA O RE? |
Através de traslocação co-traducional - são translocadas durante a sua síntese em ribossomas que estão junto á membrana do retículo. OU através de translocação pos-traducional, depois da sua tradução ter sido completada em ribossomas livres no citosol. |
|
|
EXISTEM TRÊS TEORIAS QUANTO Á VIA DE SECREÇÃO DE PROTEÍNAS. EXPLICITA CADA UMA |
Modelo da Maturação das Cisternas - Á medida que as vesículas se deslocam para o complexo de Golgi, vão se fundindo na fase cis e formam novas cisternas. Modelo das Cisternas Estáveis - São sempre as mesmas cisternas, apenas entra material diferente nestas |
|
|
EXISTEM DUAS TEORIAS QUANTO Á VIA DE SECREÇÃO DE PROTEÍNAS. EXPLICITA CADA UMA |
Modelo da Maturação das Cisternas - Á medida que as vesículas se deslocam para o complexo de Golgi, vão se fundindo na fase cis e formam novas cisternas. Modelo das Cisternas Estáveis - São sempre as mesmas cisternas, apenas entra material diferente nestas |
|
|
O QUE DETERMINA O LOCAL PARA O QUAL VAI UMA PORTEÍNA SINTETIZADA POR UM RIBOSSOMA? |
O seu péptido sinal. |
|
|
EXPLICA DETALHADAMENTE O QUE SE SUCEDE DURANTE A TRASLOCAÇÃO COTRADUCIONAL |
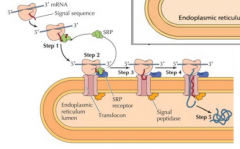
Á medida que a proteína é sintetizada pelo ribossoma uma péptido-sinal surge. Este é reconhecido pela SRP(signal recongnition particle). A SRP acompanha o complexo(ribossoma + proteína) para a membrana do retículo, onde se liga a um receptor membranar - recetor SRP. De seguida o SRP é libertado e o ribossoma liga-se a um complexo de translocação, sendo o péptido sinal inserido no complexo de tranlocação e consequentemente na membrana. Aqui recomeça a tradução e a proteína contendo o péptido sinal continua a ser traduzida para o interior da membrana. No final a peptidase sinal cliva o péptido sinal da proteína sintetizada e liberta-a para o lumen do RE. |
|
|
EXPLICA DETALHADAMENTE O QUE SE SUCEDE DURANTE A TRASLOCAÇÃO POSTRADUCIONAL |
As proteínas são sintetizadas em ribossomas livres no citosol. A proteína sintetizada é deslocada para o retículo com ajuda de chaperonas citosólicas (Hsp70), que mantém a proteína desdobrada. O péptido-sinal destas proteínas é reconhecido por duas proteínas receptoras ( complexo Sec62/63) que estão associadas ao complexo de translocação da membrana do retículo. A proteína entra pelo complexo de translocação ainda desdobrada e uma outra chaperona Hsp70(BiP) puxa a proteína para o interior do RE. |
|
|
COMO É QUE UMA PROTEÍNA INTEGRAL FICA ICORPORADA NA MEMBRANA PLASMÁTICA? |
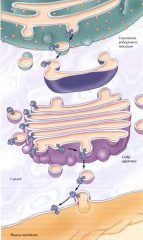
É primeiro incorporada na membrana do RE e depois segue a sequência normal RE->vesílcula->complexo de Gogi (etc) até ser inserida na membrana. Faz este percurso sempre inserida na membrana de todas as estruturas pelas quais passa. |
|
|
COMO É QUE UMA PROTEÍNA UNIPASSE É INSERIDA NA MEMBRANA DO RETÍCULO? |
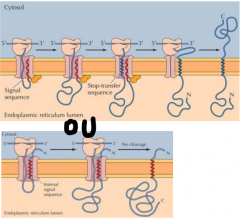
Ocorre como a tranlocação cotraducional, em que a peptidase sinal cliva o péptido sinal durante a translocação no complexo de translocação. A diferença é que há uma sequência de aminoácidos( sequência stop-traslocação) na proteína que altera o complexo de translocação e impede a translocação da proteína para o interior do retículo. Esta alteração no complexo de translocação fecha este complexo e insere a proteína na membrana. Pode continuar a tradução da proteína no citosol pelo ribossoma após a inserção. Alternativamente pode ser a sequência sinal a sequência-stop que se insere na membrana, não sendo assim clivada pela peptidase sinal. |
|
|
COMO É QUE UMA PROTEÍNA MULTIPASSE É INSERIDA NA MEMBRANA DO RETÍCULO?(DISPENSA-SE A INSERÇÃO DETALHADA) |
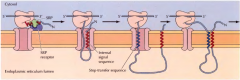
É igual á proteína unipasse mas contém mais que uma sequência-stop e várias porções desta proteína vão sendo inseridas na membrana do retículo. |
|
|
COMO É QUE UMA PROTEÍNA FICA ANCORADA AO RETÍCULO? |
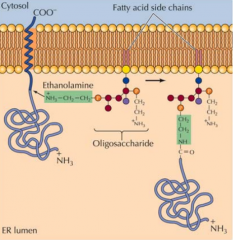
Enquanto se dá a translocação da proteína para o interior do retículo, no interior do retículo forma-se um lípido(GPI) que está unido á bicamada fosfolípidica. Quando a proteína está finalizada uma enzima cliva-a e une-a ao GPI, ficando esta associada á membrana. As proteínas depois de secretadas na membrana, ficam no exterior da membrana. |
|
|
QUE MECANISMOS EXISTEM PARA VERIFICAR A VALIDADE DAS PROTEÍNAS NO INTERIOR DO RE? |
Existem enzimas e chaperonas (calnexina e calreticulina) que controlam. |
|
|
O QUE SE SUCEDE A UMA PROTEÍNA QUE ESTÁ MAL-FORMADA NO INTERIOR DO RETÍCULO? |
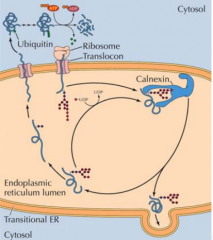
É detetada pela UDP ou por chaperonas, é enviada de novo para uma chaperona(calnexina). Se depois de vários ciclos continuar mal-formada é retro-translocada de volta para o citosol através do complexo de translocação e marcada com Ubiquitina para depois ser degradada. |
|
|
QUAL É A DIFERENÇA ENTRE PROTEOGLICANO E GLICOPROTEÍNA? |
Num proteoglicano o glícido ocupa uma percentagem maior em massa do que a proteína |
|
|
QUE NOME TEM A LÍPIDO QUE EXISTE NA MEMBRANA DO RE EM QUE É SUPORTADA A CONSTRUÇÃO DE UM OLIGOSSACARÍDEO? |
Dolicol fosfato |
|
|
RESUME A SÍNTESE DE UM OLIGOSSACARÍDEO QUE ACONTECE FORA DA MEMBRANA DO RE |
Vão sendo adicionadas unidades de N-acetilglucosamina ao dolicol fosfato, sendo a primeira adicionada com mais um grupo fosfato. É adicionada mais uma unidade de N-acetilglucosamina e depois são adicionadas 5 unidades de manose. |
|
|
RESUME A SÍNTESE DE UM OLIGOSSACARÍDEO QUE ACONTECE NO LUMEN RE |
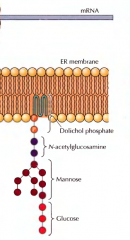
Ocorre uma translocação do oligossacarídeo do exterior para o interior do RE através de um fosfolípido - a flipase. São adicionadas mais 4 unidades de manose ao oligossacarídeo. A síntese fica completa com a adição de três unidades de glucose. |
|
|
O QUE SE SUCEDE APÓS A SÍNTESE DO OLIGOSSACARÍDEO NO LUMEN DO RE? |
Ocorre a glicosilação de uma proteína - a adição do oligossacarídeo á proteína. |
|
|
COMO SE PROCESSA A GLICOSILAÇÃO DA PROTEÍNA NO RE? |
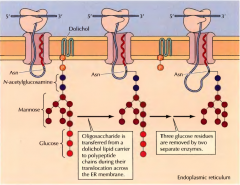
Ao mesmo tempo que se dá a translocação do oligossacarídeo para o interior do lúmen do RE e com ajuda de enzimas o oligossacarídeo é transferido no interior do retículo para uma proteína. Fica ligada a N-acetilglucosamina a uma molécula aceitadora da proteína( aparagine(Asn)). São removidas as três unidades de glucose por duas enzimas. |
|
|
SINTETIZA O QUE SE SUCEDE A UMA PROTEÍNA DEPOIS DE SER SINTETIZADA NO RE? |
Forma vesículas que se fundem para formar um intermédio entre o complexo de Golgi e o RE. As proteínas são depois transportadas para a face cis do complexo de Golgi, vão sendo transportadas pelo complexo de Golgi e são modificadas nas cisternas. São depois transportadas para a face trans do complexo de Golgi onde são selecionadas para a membrana plasmática, secreção, endossomas ou lisossomas. |
|
|
DE QUE MANEIRA SÃO MODIFICADAS AS GLICOPROTEÍNAS NO COMPLEXO DE GOLGI NO CASO DAS PROTEÍNAS QUE SE DESTINAM AOS LISOSSOMAS?(PROTEÍNAS LISOSSOMAIS). |
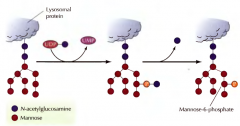
Na face cis a manose é fosofrilizada por duas enzimas( são adicionados fosfato-N-acetilglucosaminas á manose, e removida a N-acetilglucosamina). Assim fica manose-6-fosfato. Esta glicoproteína é reconhecida por um recetor de manose-6-fosfato na face trans do complexo de Golgi, que direciona o transporte destas proteínas para endossomas e lisossomas. |
|
|
COMO É QUE UMA ENZIMA SABE QUE É NECESSÁRIO ALTERAR UMA GLICOPROTEÍNA DE SERTA FORMA PARA ELA IR PARA UM LISOSSOMA? |
Possui marcadores. |
|
|
EM QUE PARTE DO RETÍCULO SÃO SINTETIZADOS LÍPIDOS? |
No RE liso |
|
|
QUE LÍPIDOS É QUE SÃO SINTETIZADOS NO RETÍCULO LISO? |
Fosfatotidilcolina, fosfatotidilletanolamina, fosfatidilserina, fosfatidilnotisol |
|
|
COMO SE PROCESSA A SÍNTESE DE FOSFOLÍPIDOS NO REL? |
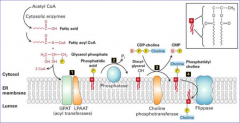
No lado exterior da membrana: Ácidos gordos são transferidos de coenzimas A(CoA) que os carregam, para o glicerol-3-fosfato através de enzimas da membrana(acyl transferases). Daqui resulta o ácido fosfatídico que é inserido na membrana. De seguida enzima fosfatase converte o ácido fosfatídico em diacilglicerol. Depois a enzima colina fosfotransferase altera o diacilglicerol em fosfatotidilcolina, fosfatotidilletanolamina, fosfatidilserina, fosfatidilnotisol e a flipase coloca os fosfolípidos no interior do lúmen do RE. |
|
|
EM QUE CÉLULAS É MAIS ABUNDANTE O RETÍCULO LISO? |
Células ativas no metabolismo de lípidos. |
|
|
ONDE É SINTETIZADA A CERAMIDA? |
No retículo liso. |
|
|
QUE LÍPIDOS PODE ORIGINAR A MODIFICAÇÃO DA CERAMIDA? |
Esfingomielina ou glicolípidos. |
|
|
ONDE SE FORMA A ESFINGOMIELINA? |
No complexo de Golgi. |
|
|
COMO SE FORMA A ESFINGOMIELINA NO COMPLEXO DE GOLGI? |
É sintetizada apartir da adição de fosforilcolina da fosfatidilcolina para a ceramida no lúmen do complexo de Golgi |
|
|
COMO SE FORMAM GLICOLÍPIDOS NO COMPLEXO DE GOLGI? |
Por adição de glícidos á ceramida. |
|
|
COMO SE FORMAM GLICOSILCERAMIDA NO COMPLEXO DE GOLGI? |
Por adição de glucose á esfigomielina do lado citosólico(exterior da membrana) |
|
|
PORQUE É QUE OS GLICOLíPIDOS ESTÃO SEMPRE DO LADO EXTERIOR DAS MEMBRANAS? |
Pois quando são sintetizados no interior do complexo de Golgi, não são capazes de se translocar para o exterior da membrana. Assim depois de serem transportados por vesículas e de se fundirem com a membrana plasmática, ficam do lado de fora. |
|
|
COMO É QUE FOSFOLÍPIDOS SINTETIZADOS NO RE SÃO TRANSFERIDOS DIRETAMENTE PARA ORGANELOS COMO A MITOCÔNDRIA? |
Através de proteínas de transferência de lípidos. |
|
|
QUE MANEIRAS TÊM OS LIPIDOS DE SE TRANSFERIR ENTRE MEMBRANAS? |
Difusão de monómeros, colisão ativada, através de proteínas, formação de condutas hidrofóbicas, disturbio e difusão, ativação e colisão, fusão. |
|
|
EM QUE CONSISTE A TRANSFERÊNCIA DE LÍPIDOS ENTRE DUAS MEMBRANAS POR COLISÃO ATIVADA? |
Durante a colisão de duas membranas um lípido é ativado e passa para outra membrana. |
|
|
COMO SÃO TRANPORTADAS AS PROTEÍNAS E LÍPIDOS DO RE PARA O COMPLEXO DE GOLGI? |
Através de vesículas de transporte |
|
|
COMO É A ORIENTAÇÃO DE PROTEÍNAS MEMBRANARES NO RE E NO COMPLEXO DE GOLGI? |
A orientação é igual nos dois. Se a proteína tiver como destino fica do lado citosolico da membrana, vai estar na parte interior do RE e do complexo de Golgi. Assim como o contrário. |
|
|
O QUE DETERMINA SE UMA PROTEÍNA VOLTE PARA O COMPLEXO DE GOLGI? |
O facto de possuir uma sequência sinal. |
|
|
QUE SEQUÊNCIA DE AMINOÁCIDOS POSSUEM AS PROTEÍNAS QUE SE DESTINAM A VOLTAR PARA O LUMEN DO RE? |
KDEL - Lis-Asp-Glu-Leu |
|
|
QUE SEQUÊNCIA DE AMINOÁCIDOS POSSUEM AS PROTEÍNAS MEMBRANARES DO RE? |
KKXX - Lis-Lis |
|
|
O QUE DETERMINA QUE AS PROTEÍNAS ANCORADAS SEJAM EXPORTADAS DO RE? |
O facto de estarem ligadas ao GPI |
|
|
QUE SEQUÊNCIAS DE AMINOÁCIDOS POSSUEM AS PROTEÍNAS TRANSMEMBRANARES QUE AS SINALIZAM PARA SEREM EXPORTADAS? |
Sequências di-acidicas (Asp-Asp ou Glu-Glu); sequências di-hidrofóbicas (Met-Met). |
|
|
QUE IMPORTÂNCIA TÊM AS SEQUÊNCIAS DE AMINOÁCIDOS KDEL E KKXX? |
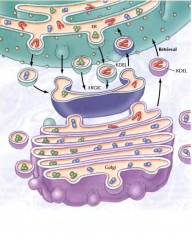
Permite que quando estão em vesículas de transporte para o complexo de Golgi, sejam recicladas(recuperação seletiva de proteínas do RE), no complexo intermedio(CIRG) ou no complexo de Golgi. |
|
|
QUE MANEIRAS TÊM AS PROTEÍNAS E OS LÍPIDOS DE SER TRANSLOCADOS DO COMPLEXO DE GOLGI? |
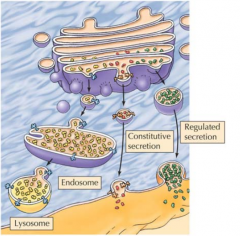
Através de secreção constitutiva, regulada ou transporte seletivo para os lisossomas. |
|
|
DE QUE FORMA EXISTEM PROTEÍNAS NO COMPLEXO DE GOLGI? |
Apenas associadas á membrana, não existem livres no lúmen. |
|
|
O QUE DIFERE NO PROCESSO EM QUE AS PROTEÍNAS FICAM RETIDAS NO RE E NO COMPLEXO DE GOLGI? |
As que ficam retidas no RE são recicladas e enviadas de volta. As que ficam no complexo de Golgi possuem sequências de aminoácidos que determinam que estas não saem. |
|
|
EM QUE CONSISTE A SECREÇÃO CONSTITUTIVA DO COMPLEXO DE GOLGI? |
É a secreção continua e não regulada de proteínas e lípidos, tanto para integrarem a membrana plasmática como para serem secretados para fora da célula. |
|
|
EM QUE CONSISTE A SECREÇÃO REGULADA DO COMPLEXO DE GOLGI? |
É a secreção de proteínas e lípidos específicos em resposta a sinais ambientais. |
|
|
DÁ 3 EXEMPLOS DE SECREÇÃO REGULADA DO COMPLEXO DE GOLGI |
Libertação de hormonas, libertação de neurotransmissores, libertação de enzimas digestivas. |
|
|
COMO É O MECANISMO DA SECREÇÃO REGULADA DO COMPLEXO DE GOLGI? |
As proteínas e lipidos são selecionadas para secreção regulada na face trans do complexo de Golgi e são colocadas em vesículas secretoras especializadas. Estas vesículas ainda imaturas, que são maiores que a vesículas de transporte, processas os seus contéudos e fundem-se umas com as outras podendo ter de aguardar um sinal biológica para se poderem fundir com a membrana. |
|
|
COMO SE DÁ O TRANSPORTE SELETIVO DE PROTEÍNAS E LÍPIDOS DO COMPLEXO DE GOLGI PARA OS LISOSSOMAS? |
As proteínas destinadas aos lisossomas são marcadas pela manose-6-fosfato, e reconhecidas por um recetor da membrana da face trans do complexo de Golgi, que as coloca em vesículas de transporte destinadas aos lisossomas. |
|
|
O QUE SÃO VESÍCULAS REVESTIDAS? |
São vesículas de transporte cobertas com por proteínas citosólicas de revestimento. |
|
|
COMO SE FORMAM AS VESÍCULAS REVESTIDAS? |
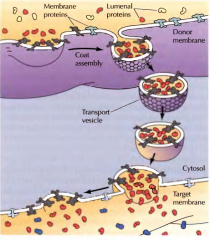
Formam-se enquanto a vesícula se separa da membrana de onde provém. A formação do revestimento dá-se com ajuda de proteínas-de- junção-GTP. |
|
|
QUAIS SÃO OS TRÊS TIPOS DE VESÍCULAS REVESTIDAS? |
COP(coat protein): - COPI; COPII. E Vesículas revestidas de clatrina. |
|
|
PARA ONDE LEVAM AS PROTEÍNAS SECRETORAS AS VESÍCULAS REVESTIDAS COP I? |
Levam entre locais dentro do complexo de Golgi. Por exemplo da face trans para a face cis. Ou se forem marcadas por KDEL e KKXX levam de volta da face cis do complexo Golgi para o RE. |
|
|
PARA ONDE LEVAM AS PROTEÍNAS SECRETORAS AS VESÍCULAS REVESTIDAS COP II? |
Levam do RE para a face cis do complexo de Golgi. |
|
|
QUAL É O PAPEL DAS VESÍCULAS REVESTIDAS COM CLATRINA? |
Promovem a endocitose de moléculas extracelulares e o transporte de moléculas da face trans do complexo de Golgi para endossomas, lisossomas ou para a membrana plasmática. |
|
|
COMO OCORRE A FORMAÇÃO DE VESÍCULAS REVESTIDAS DE CLATRINA? |

Primeiro a proteína ARF1-GDP liga-se a proteínas na membrana do complexo de Golgi e há a troca de GDP por GTP. O complexo ARF/GTP inicia a formação da vesícula recrutando proteínas adaptadoras como a GGA, que servem de locais de ligação para recetores transmembranares e para a clatrina. A clatrina tem uma função estrutural na formação da vesícula, ao distorcer a membrana e iniciar a formação da vesícula. |
|
|
O QUE SE SUCEDE APÓS A FORMAÇÃO DAS VESÍCULAS REVESTIDAS DE CLATRINA? |
A GTP é hidrolizada a GDP e o complexo ARF/GDP é libertado da membrana de forma a ser reciclado. É libertação enfraquece a ligação da clatrina á membrana e permite a enzimas e proteínas chaperonas a desintegração do revestimento. |
|
|
COMO CHEGAM AS PROTEÍNAS AOS LISOSSOMAS? |
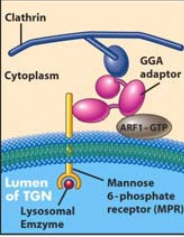
As proteínas destinadas aos lisossomas são marcadas por manose-6-fosfato. Estas proteínas ligam-se a recetores manose-6-fosfato na face trans do complexo de Golgi, que por sua vez se ligam á clatrina levando á formação das vesículas |
|
|
O QUE É A HIPÓTESE DE SNARE? |
É a hipótese que explica a fusão de vesículas através da interação entre pares específicos de proteínas transmembranares chamadas de SNAREs. |
|
|
EXPLICA COMO SE PROCEDE A FUSÃO DE VESÍCULAS DE ACORDO COM A HIPÓTESE DE SNARE |

Primeiro pequenas proteínas onde se liga GTP, as Rabs, interagem com proteínas efetoras e com v-SNARES formando o complexo de pré-fusão. Outra proteína Rab na membrana em que se vai fundir a vesícula organiza outras proteínas efetoras e t-SNARES. Quando a vesícula de transporte contacta com a membrana as proteínas efetoras ligam as membranas e dá-se a hidrólise do Rab/GTP que permite o contacto entre as v-SNARES e as t-SNARES, que aproximam as duas membranas criando instabilidade e fusão. Após a fusão dá-se o desmatelamento dos complexos SNAREs com hidrólise de ATP. |
|
|
QUE IMPORTÂNCIA TEM O CÁLCIO NA SECREÇÃO DE PROTEÍNAS EM VESÍCULAS? |
Tem importância na secreção regulada da insulina e de neurotransmissores. Nos dois casos, é a entrada de cálcio no interior da membrana que desencadeia a secreção das moléculas. |
|
|
O QUE SÃO OS LISOSSOMAS? |
São organelos celulares com membranas que possuem enzimas capazes de degradar vários compostos biológicos. São o sistema digestivo da célula. |
|
|
PORQUE CONTÉEM OS LISOSSOMAS GRANDE VARIEDADE DE ENZIMAS? |
Pois existem uma grande quantidade de substâncias que estes necessitam de degradar. |
|
|
EM QUE CATEGORIA SE INSEREM A MAIOR PARTE DAS ENZIMAS LISOSSOMAIS? PORQUÊ? |
São hidrolases ácidas, pois só estão ativas no pH ácido(5). |
|
|
PORQUE RAZÃO AS ENZIMAS LISOSSOMAIS SÓ FUNCIONAM A UM PH ÁCIDO? |
É uma medida de segurança. Caso os lisossomas se rompam, as enzimas no pH neutro do citosol deixam de funcionar e não podem degradar a célula. |
|
|
COMO MANTÉEM OS LISOSSOMAS O PH ÁCIDO NO SEU INTERIOR? |
Pois possuem uma bomba de protões na sua membrana lisossomal. Esta bomba hidrolisa o ATP e transfere protões para o interior do lisossoma, mantedo a concentração 100x superior á exterior. |
|
|
COMO SE FORMAM OS LISOSSOMAS? |
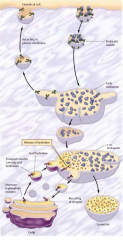
Quando vesículas de transporte da face trans do complexo de Golgi se fundem com endossomas. Ao mesmo tempo material do exterior da célula é endocitado em vesículas endociticas revestidas de clatrina e se funde com endossomas imaturos. Este acabam por originar os lisossomas. |
|
|
QUE IMPORTÂNCIA TEM O ABAIXAMENTO DO PH DURANTE A FORMAÇÃO DE LISOSSOMAS? |
O pH baixo no interior dos lisossomas faz com que as hidrolazes acidicas se dissociem do recetor de manose-6-fosfato e sejam libertadas para o interior do lisossoma onde podem assumir a sua função de digestão. |
|
|
QUAL É A DIFERENÇA ENTRE FAGOCITOSE E AUTOFAGIA? |
Na fagocitose, a célula ingere partículas, bactérias ou outras células envelhecidas, e estas são digeridas por lisossomas. Na autofagia são degradados os próprios componentes da célula por lisossomas, este processo ao contrário da fagocitose, ocorre em todas as células. |
|
|
O QUE CAUSA A DOENÇA DE GAUCHER? |
É uma falha enzima lisossomal glucocerebrosidase, que faz a hidrólise da glucocerebroside em glucose e ceramida. Este mau funcionamento da enzima provoca um mau funcionamento dos lisossomas a acumulação de glicolípidos nas células e um aumento de lisossomas para os degradar o que provoca mau funcionamento celular. |
|
|
QUE FUNÇÕES ASSUMEM OS VACÚOLOS NAS CÉLULAS VEGETAIS? |
Armazenamento, turgidez celular, regulação do pH da célula, transporte de proteínas membranares no tonoplasto. |
|
|
COMO CHEGAM AS PROTEÍNAS AO VACÚOLO VEGETAL? |
São sintetizadas no RE rugoso, transportadas pelo complexo de Golgi e selecionadas, pois possuem sequências peptidicas especiais, sendo enviadas para o vacúolo. |
|
|
QUE TRÊS FORMAS DE ENDOCITOSE EXISTEM? |
Fagocitose, pinocitose e endocitose mediada por recetores. |
|
|
EM QUE CONSISTE A FAGOCITOSE? |
Uma partícula liga-se a recetores na célula fagocitica que desencadeia a extenção de pseudópodes (regulados por actina) que englobam o material, fundindo-se as membranas e envolvendo uma material por uma membrana. Este depois é levado até um lisossoma, funde-se formando um fagolisossoma e digere o contéudo. |
|
|
COMO SE DÁ A ENDOCITOSE MEDIADA POR RECETORES (CLATRINA)? |
As moléculas que vão ser endocitadas ligam-se a recetores. Estes recetores estão concentrados em zonas cobertas de clatrina. Com ajuda da proteína diamina estas zonas começam a formar pequenas vesículas cobertas de clatrina. Estas vesículas contém os recetores e os respetivos ligandos. As vesículas depois fundem-se com endossomas e a sua cobertura de clatrina e os seus recetores são retirados |
|
|
COMO SE DÁ A ENDOCITOSE MEDIADA POR RECETORES (CAVEOLINA)? |
As moléculas ligam-se a caveolas, pequenas invaginações na membrana que contéem caveolina, que funciona como recetor, ativando a endocitose. |
|
|
COMO SE DÁ A MACROPINOCITOSE? |
Não é depende de recetores e engloba de imediato líquidos. |
|
|
O QUE SÃO CORPOS MULTI-VESICULARES? |
São endossomas com contéudos ainda nao digeridos(outras vesículas), que ainda não maturaram para lisossomas. |
|
|
COMO É O COLESTEROL INTERGRADO NAS CÉLULAS? |
O colesterol é transportado pela corrente sanguínea através de proteínas de baixa densidade (LDL). Para o colesterol entrar numa célula é necessário que a LDL se ligue a recetores específicos membranares nas zonas cobertas com clatrina e seja endocitado. O recetor é depois reciclado e a LDL segue para os lisossomas onde o colesterol é libertado para o uso da célula. |
|
|
O QUE É A HIPERCOLESTEROLÉMIA FAMILIAR? |
É uma doença causada por uma mutação no recetor de LDL, o que impede a endocitose de colesterol e provoca um aumento de colesterol na corrente sanguínea. |
|
|
QUAIS SÃO OS ELEMENTOS FILAMENTOSOS DO CITOESQUELETO? |
Filamentos intermédios, filamentos de actina, microtubulos. |
|
|
QUAIS SÃO AS PROTEÍNAS MOTORAS QUE CONSTITUEM O CITOESQUELETO? |
Miosinas, Cinesinas e dineínas. |
|
|
QUAIS SÃO AS FUNÇÕES DO CITOESQUELETO? |
Dão estrtura e suporte; promovem o transporte intracelular; permitem a contractabilidade e mobilidade de componentes, promovem a organização espacial. |
|
|
O QUE É A ACTINA? |
É uma proteína que forma microfilamentos. |
|
|
ONDE SÃO MAIS ABUNDANTES OS MICROFILAMENTOS DE ACTINA? |
Debaixo da membrana plasmática, dando suporte mecânico e permitindo movimento membranar. |
|
|
A ACTINA ASSUE DUAS FORMAS. QUAIS SÃO? |
Actina G - monomérica que está associada sempre a trios( cada actinaG está associada a mais duas), formando actina filamentosa ActinaF. |
|
|
PORQUE TÊM OS FILAMENTOS DE ACTINA POLARIDADE? QUAL A SUA IMPORTÂNCIA? |
Devido à orientaçã da actinaG, os monómeros ficam todos no mesmo sentido, assim como os grupos fosfato do ATP, o que faz com que os microfilamentos de actina adquiram polaridade. Esta polaridade é importante para a forma como se agrupam os monómero e para a direção do movimento da miosina |
|
|
COMO É QUE OS FILAMENTOS DE ACTINA SE POLIMERIZAM? |
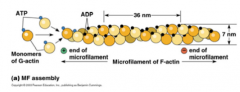
Primeiro dá-se a nucleação, em que se formam agregados de três monómeros de actina. Os filamentos de actina depois crescem através da adição de monómeros ás duas extremidades , mas a extremidade positiva cresce 10 vezes mais rápido que a extremidade negativa. Os monómeros de actina também unem ATP que apesar de não ser necessário para a polimerização, é o que determina a rapidez com os monómeros se polimerizam. Os monómeros com ADP polimerizam mais lentamente |
|
|
QUE ESTRUTURAS PODEM FORMAR OS FILAMENTOS DE ACTINA? |
Amontoados(bundle) de actina ou redes de actina. |
|
|
QUE FUNÇÃO TEM O TIPO DE AMONTOADO DE ACTINA QUE É CONSTITUIDO POR FILAMENTOS PRÓXIMOS ALINHADOS EM PARALELO? QUE PROTEÍNA ESTÁ ENVOLVIDA NA FORMAÇÃO DESTAS ESTRUTURAS? |
Suporta projeções da membrana plasmática como microvilosidades. È a fimbrina. |
|
|
QUE FUNÇÃO DESEMPENHA A FIMBRINA NOS FILAMENTOS DE ACTINA? |
Promove a estabilidade dos filamentos, ao agrupar dois filamentos paralelos de forma a estarem mais próximos. |
|
|
QUE DIFERENÇA POSSUEM AS LIGAÇÕES FEITAS POR ALFA-ACTININA FACE ÁS LIGAÇÕES FEITAS POR FIMBRINA? |
As ligações feitas por alfa-actinina mantêm os filamentos mais afastados permitindo a passagem da proteína motora miosina que impede a contração dos filamentos. |
|
|
QUE FUNÇÃO TEM A FILAMINA NAS REDES DE ACTINA? QUE CARACTERÍSTICAS TEM A PROTEÍNA FILAMINA? |
A filamina é composta por duas sub-unidades que se ligam a filamentos diferentes na rede. É esta proteína que permite a ligação entre filamentos de actina na rede e a estabilização desta conformação. |
|
|
DE QUE FORMA INFLUENCIAM A MEMBRANA PLASMÁTICA OS FILAMENTOS DE ACTINA? |
Unem-se a proteínas membranares, determinam a estrutura, dão suporte à membrana e estão envolvidos em movimentos da membrana plasmática.. |
|
|
EM QUE INTERAÇÕES ENTRE CÉLULAS ESTÃO PRESENTES OS FILAMENTOS DE ACTINA? |
Nas junções de aderência. |
|
|
QUE FUNÇÃO TÊM OS FILAMENTOS DE ACTINA, EM JUNÇÕES DE ADERÊNCIA, NAS CÉLULAS EPITELIAIS? |
Formam uma estrtura que une as células, para se manterem próximas. |
|
|
COMO SÃO COMPOSTAS AS JUNÇÕES DE ADERÊNCIA NAS CÉLULAS EPITELIAIS? |
São unidas por proteínas transmembranares, as caderinas que possuem uma parte fora da célula e que promovem a união formando complexos, as catetinas, que se associam aos filamentos de actina no interior das células. |
|
|
COMO SE FORMAM AS MICROVILOSIDADES? |
Através de extensões de filamentos de actina que promovem o prolongamento da membrana plasmática. Os filamentos de actina são unidos pelas proteínas fimbrina e por vilina. Na base as microvilosidades os filamentos de actina unem-se a proteínas o que estabiliza as microvilosidades. |
|
|
COMO É QUE OS FILAMENTOS DE ACTINA INTERVÊEM EM FUNÇÕES ALIMENTARES E DE LOCOMOÇÃO? |
São responsáveis pela formação de pseudópodes. |
|
|
O QUE É A MIOSINA? QUAL É A SUA IMPORTÂNCIA? |
É uma proteína que converte ATP em energia mecânica e gera movimento dos filamentos de actina. É importante na contração muscular. |
|
|
QUANDO HÁ MOVIMENTO DOS FILAMENTOS DE ACTINA NUM SENTIDO, QUAL É A POLARIDADE DOS FILAMENTOS DE ACTINA NESSE SENTIDO?PORQUÊ? |
Têm polaridade positiva, pois é o lado positivo dos filamentos que se polimeriza mais rapidamente, logo é o que segue á frente. |
|
|
COMO É COMPOSTO A MAIOR PARTE DO CITOPLASMA DAS CÉLULAS QUE CONSTITUEM AS FIBRAS MUSCULARES? |
Por miofibrilhas, que são associações entre filamentos de miosina e de actina. |
|
|
COMO ESTÃO DIVIDIDAS AS MIOFIBRILHAS? |
Em cadeias de unidades contracteis sarcomeros, que são responsáveis pelo aspeto estriado do musculo estriado e cardiaco. |
|
|
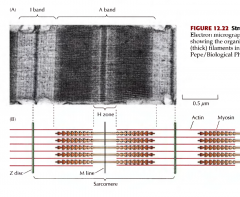
DESCREVE A CONSTITUIÇÃO DOS SARCÓMEROS E AS DIEFRENTES CARACTERÍSTICAS |
São delimitados por linhas, linhas Z. Os sarcómeros são compostos por zonas mais escuras, bandas A, e zonas mais claras, bandas I. As bandas I são mais claras pois possuem apenas finos filamentos de actina, as bandas A possuem espessos filamentos de miosina e filamentos de actina. A zona H, no meio das bandas A apenas possui miosina. Os filamentos de actina estão unidos ao disco Z por proteínas alfa-actinina. Os filamentos de miosina estão unidos á linha M(no meio da zona H na banda A) |
|
|
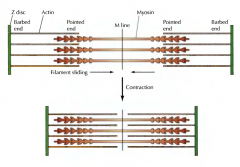
O QUE SE SUCEDE DURANTE A CONTRAÇÃO MUSCULAR? |
A zona I e a zona H desaparecem quase por completo. Isto resulta de uma interação entre os filamentos de miosina e de actina. Os filamentos de miosina provocam o movimento contrário dos filamentos de actina face aos filamentos de miosina , que ficam acumulados na banda A e na zona H, ficando o músculo contraído. Neste processo não há alteração da banda A. |
|
|
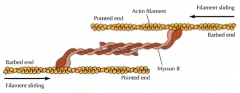
COMO SE DÁ O MOVIMENTO DA MIOSINA AO LONGO DA ACTINA, PROMOVENDO A CONTRAÇÃO MUSCULAR? |
A ligação do ATP à miosina, providencia a energia necessária ao movimento da miosina e promove a dissociação da actina á miosina. A hidrólise de ATP depois provoca alterações na proteína que a faz ligar de novo, mas mais a frente, ao filamento de actina, sendo libertado ADP e fosfato(P). |
|
|
COMO É QUE A CONTRAÇÃO ACTINA-MIOSINA PROMOVE A CITOCINESE? |
No fim da mitose, um anel de contração é formado de baixo da membrana plasmática, que constiste em filamentos de actina unidos uns aos outros por miosina. Á medida que o anel de contração se encurta os filamentos de actina também se vão encurtando( por despolimerização). É o encurtamento dos filamentos de actina que promove a divisão da célula. |
|
|
COMO SÃO FEITOS OS MOVIMENTOS CITOPLASMÁTICOS EM CÉLULAS VEGETAIS? |
A miosina liga os organelos aos filamentos de actina. À medida que a miosina se move ao longo dos filamentos de actina, também se movem os organelos. |
|
|
PORQUE TÊM OS FILAMENTOS INTERMÉDIOS, O NOME INTERMÉDIOS? |
Pois têm diametro intermédio entre os filamentos de actina(menor) e os microtubulos(maior) |
|
|
QUE FUNÇÃO DESEMPENHAM OS FILAMENTOS INTERMÉDIOS? |
Apenas função estrututral, dando suporte mecânico a células e tecidos. |
|
|
QUE TIPOS DE PROTEÍNAS EXISTEM NOS FILAMENTOS INTERMÉDIOS? EM QUE CÉLULAS DESEMPENHAM FUNÇÕES? |
Tipo I e II, são keratinas ácidas e básicas respetivamente e existem nas células epiteliais pelo menos um tipo básico e um ácido. Tipo III: vimentina(fibroblastos), desmina(células musculares), proteínas da glia, periferina( neurónios periferais). Tipo IV: proteínas neurofilamentosas, nos neurónios. Tipo V, lamininas nucleares, na lâmina nuclear. Tipo VI: nestina, em células estaminais dos sistema nervoso. |
|
|
QUE KERATINAS CONSTITUEM O CABELO E AS UNHAS? |
Keratinas tipo II, keratinas básicas. |
|
|
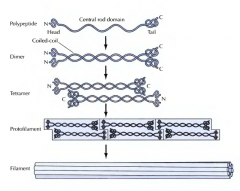
COMO SE FORMA UM FILAMENTO INTERMÉDIO? |
Duas cadeias de polipeptidos de helice-alfa entrelaçam-se para formar dimeros, que se juntam antiparalelamente(N com C) para formar tetrameros, associados uns com os outros formam protofilamentos que em grupos de 8 formam filamentos. |
|
|
COMO SE ORIENTAM OS FILAMENTOS INTERMÉDIOS NO INTERIOR DA CÉLULA DE FORMA A CONSTITUIR O CITOESQUELETO? |
Formam um anel em rede no citoplasma, em torno do núcleo. A keratina e a vimentina fixam-se ao invólucro nuclear fixando a posição do núcleo no interior da célula. Os filamentos intermédios estão também associados aos microtúbulos e aos filamentos de actina formando o citoesqueleto. |
|
|
QUE TIPO DE JUNÇÕES INTERCELULARES FORMAM OS FILAMENTOS INTERMÉDIOS DE KERATINA NAS CÉLULAS EPITELIAIS? |
Desmossomas e hemidesmossomas. |
|
|
O QUE SÃO OS DESMOSSOMAS? |
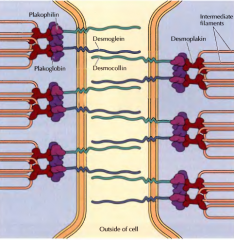
Desmossomas são junções entre células adjacentes. Nestas junções os filamentos intermédios estão no interior da célula, associados a proteínas intracelulares, as desmoplaquinas, através de keratina. As desmoplaquinas estão associadas a proteínas transmembranares que promovem o contacto entre células, as caderinas(desmoglina e desmocolina). |
|
|
O QUE SÃO HEMIDESMOSSOMAS? |
São junções em que os filamentos intermédios estão ligados dentro da célula através de plectina a integrinas que promovem o contacto com a matriz extracelular( ao contrário dos desmossomas que contactam com outras células) |
|
|
QUE FUNÇÃO IMPORTANTE TÊM OS DESMOSSOMAS E OS HEMIDESMOSSOMAS? |
Dão estabilidade ás células, fixando-as. |
|
|
O QUE É,COMO É CONSTITUIDA E QUE IMPORTÂNCIA TEM A LÂMINA NUCLEAR? |
É uma rede fibrilhar no interior do núcleo, constituida por filamentos intermédios e proteínas associadas ás membranas nucleares. Desempenha funções inportantes na replicação do DNA, na divisão celular, organiza a cromatina e fixa os poros nucleares num envólucro nuclear. |
|
|
QUAL É A ÚNICA PROTEÍNA QUE CONSTITUI OS MICROTÚBULOS? |
É a tubulina, proteína globular. |
|
|
QUE DOIS POLIPÉPTIDOS SÃO CONSTITUIDOS POR TUBULINA E CONSTITUEM OS MICROTÚBULOS? |
Tubulina-alfa e tubulina-beta. |
|
|
ONDE SURGEM MICROTÚBULOS EM MAIOR QUANTIDADE? QUE POLIPÉPTIDO POSSUEM A MAIS NESTA ZONA? |
Junto ao centrossoma, possuem tubulina-gama |
|
|
COMO É A ESTRUTURA GERAL DOS MICROTÚBULOS? |
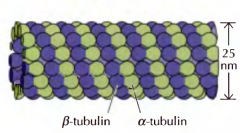
São constituidos por tubulinas-alfa e beta aos pares que formam filamentos paralelos enrolados em torno de poro sem nada. |
|
|
QUE CONSEQUÊNCIA TEM NOS MICROTÚBULOS O ARRANJO DA ALFA-TUBULINA E BETA-TUBULINA PARALELAMENTE QUE SE ASSEMELHA AOS FILAMENTOS DE ACTINA? |
A polaridade |
|
|
QUE IMPORTÂNCIA TEM A POLARIDADE DOS MICROTÚBULOS?
|
É importantes para o seu movimento. |
|
|
QUE OUTRAS SEMELHANÇAS TÊM OS MICROTÚBULOS COM OS FILAMENTOS DE ACTINA, PARA ALÉM DA POLARIDADE? |
A extremidade negativa cresce menos rapidamente que a extremidade positiva. Os microtúbulos podem-se desmantelar e voltar a construir-se polimerizando e despolimerizando a tubulina, tal como é feito nos filamentos de actina. A tubulina também possui GTP que desepenha uma função semelhante ao ATP nos filamentos de actina, favorecendo a despolimerização |
|
|
COMO É CONHECIDA A PROPRIEDADE DOS MICROTÚBULOS QUE PERMITE A SUA POLIMERIZAÇÃO E DESPOLIMERIZAÇÃO? |
Instabilidade dinâmica. |
|
|
COMO É CONTROLADA A INSTABILIDADE DINÂMICA DOS MICROTÚBULOS? |
Através de proteínas as MAPs(Microtubule associated protein) |
|
|
QUE FUNÇÕES ASSUMEM AS MAPs PARA CONTROLAR A INSTABILIDADE DINÂMICA DOS MICROTÚBULOS? |
Estabilizam os microtúbulos aos cerrar as suas extremidades, outras desfazem os microtúbulos, outras guiam microtúbulos para locais específicos. |
|
|
NAS CÉLULAS ANIMAIS, APARTIR DE QUE ESTRUTURA SE EXTENDEM OS MICROTÚBULOS? |
Apartir do centrossoma. |
|
|
COMO ESTÃO ORGANIZADOS OS MICROTÚBULOS APARTIR DO CENTROSSOMA DURANTE A INTERFASE? |
As extremidades negativas situam-se junto ao centrossoma, que está localizado junto do núcleo e extendem-se até á periferia da célula. É apartir do centrosso que se comessam a formar os microtúbulos. |
|
|
QUE PROTEÍNA É NECESSÁRIA ESTAR NUM LOCAL PARA AÍ COMEÇAR A FORMAÇÃO DE MICROTÚBULOS? |
A tubulina-gama. |
|
|
ONDE EXISTEM, COMO SÃO CONSTITUÍDOS E QUAL A FUNÇÃO DOS CENTRÍOLOS? |

Os centríolos existem nos centrossomas, são constituidos por 9 tripletos de microtúbulos e são necessários como base. |
|
|
QUAIS SÃO AS PROTEÍNAS MOTORAS RESPONSÁVEIS PELO MOVIMENTO DOS MICROTÚBULOS? |
Cinesinas e dineínas |
|
|
QUE OUTRA PROTEÍNA EXISTE NOS CENTRÍOLOS PARA ALÉM DAS TUBULINAS ALFA, GAMA, BETA? |
Tubulina delta. |
|
|
A CINEZINA E A DINEÍNA MOVEM-SE EM SENTIDOS OPOSTOS AO LONGO DOS MICROTÚBULOS. EM QUE SENTIDO SE MOVE CADA UMA? |
Dineína para o negativo e cinesina para o positivo. |
|
|
COMO É COMPOSTA A CINESINA? QUE FUNÇÃO TEM? |
Por duas sub-unidades enroladas uma na outra. Transporta elementos do citoplasma no sentido oposto ao núcleo(para a periferia). As partes(2) da cinesina que contactam com os microtubulos ligam-se a estes e ao ATP, hidrolizam-do e obtendo a energia necessária para o seu movimento. |
|
|
COMO É COMPOSTA A DINENINA? QUE FUNÇÃO TEM? |
Por três cadeias pesadas complexadas com várias cadeias leves. As cadeias pesadas formam zonas que se ligam aos microtubulos e hidrolizam o ATP fornecendo a energia necessária para o seu movimento.Transporta coisas da periferia para o centro da célula com ajuda de uma proteína, a dinactina. |
|
|
NAS CÉLULAS VEGETAIS NÃO HÁ CENTROSSOMAS. COMO SE ORGANIZAM OS MICROTUBULOS? |
Apartir de acumulações de tubulina gama. |
|
|
O QUE EXISTE NA BASE DOS FLAGELOS QUE TAMBÉM EXISTE NOS MICROTUBULOS? |
Tubulina gama. |
|
|
EM QUE DIFEREM OS FLAGELOS E OS CÍLIOS? |
Os cílios existem em grande número, os flagelos normalmente não são mais de dois por célula. Os flagelos são mais longos e movem-se num movimento ondulante, enquantos os cílios de movem no movimento cordenado para a frente e para trás. Os flagelos têm função locomotiva, enquanto o cílios podem também ter funções alimentares ou de limpeza. |
|
|
COMO É A ESTRUTURA DOS CÍLIOS E DOS FLAGELOS? |
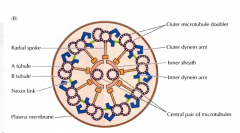
È o axonema. É composto por nove pares de microtubulos em anel rodeando um par de microtubulos. Os pares do anel estão fundidos, cendo compostos por um tubulo A e um tubulo B. Os pares do anel radial estão unidos ao anel central e unidos entre si por nexina. Estão ligadas também dineinas ao tubulos A e é a sua atividade motora que permite o movimento dos cilios e dos flagelos. |
|
|
COMO ESTÃO FIXOS OS CÍLIOS E OS FLAGELOS ÁS CÉLULAS? |
Pela parte negativa dos microtubulos ao corpo basal. |
|
|
EXPLICA DETALHADAMENTE COMO SE DÁ O MOVIMENTO DOS CÍLIOS E DOS FLAGELOS |
As dieninas que ligam os microtubulos A aos B deslizam sob os microubulos B em direção á parte negativa fazendo com que estes se curvem o que dá origem ao seu movimento. |
|
|
DURANTE A PROFASE, NA MITOSE, OS MICROTUBULOS DESOSGANIZAM-SE. O QUE FORMAM? |
Formam o fuso acromático. |
|
|
O QUE SE SUCEDE NO ÍNICIO DA MITOSE AOS MICROTÚBULOS? |
Quando começa a mitose os centríolos e os centrossomas já estão duplicados. Estes separam-se para os pólos, formando os dois pólos do fuso acromático. Com a célula a entrar em mitose aumenta exponencialmente a despolimerização dos microtúbulos(ficando mais curtos) e o número de microtúbulos. |
|
|
QUE TIPOS DE MICROTUBULOS ENVOLVIDOS NA MEIOSE EXISTEM? |
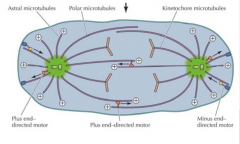
Microtubulos do cinetocoro que se ligam aos centrómeros dos cromossomas, associando-se a proteínas e formando o cinetócoro. Microtubulos cromossomais que se ligam ás extremidades dos cromossomas. Microtúbulos polares que estão ligados entre si e não aos cromossomas. Microtubulos astrais que se extendem dos cromossomas para a periferia da célula tendo as pontas positivas livres. |
|
|
COMO SE DÁ A SEPARAÇÃO DOS CROMOSSOMAS DURANTE A MITOSE? |
Os cromossomas movem-se para pólos opostos devido a movimentos da anafase A que consitem na despolimerização dos microtubulos nas partes negativas e do puxar dos cinetocoros no sentido dos microtúbulos. Na anafase B omovimento em sentidos opostos de microtubulos promovido por proteínas que se prendem a microtubulos opostos e se movem no sentido positivo dos microtubulos. Ao mesmo tempo durante a anafase B os microtubulos astrais fixam-se à perfiferia e puxam os polos do fuso acromático. Estas forças opostas promove a separação dos cromossomas. |
|
|
O QUE É O CENTRÓMERO? |
Zona do cromossoma com cromatina mais condensada onde está o cinetocoro. |
|
|
O QUE É O CINETOCORO? |
Estrutura proteica dentro junto aos centrómeros onde os microtúbulos do fuso acromático se ligam. |
|
|
QUE TIPOS DE JUNÇÕES SE FORMAM ENTRE AS INTEGRINAS E MATRIZ EXTRACELULAR? |
Hemidesossomas e junções de adesões focais. |
|
|
COMO E O QUE LIGAM AS JUNÇÕES DE ADESÕES FOCAIS DA MATRIZ EXTRACELULAR ÁS CÉLULAS? |
Estas junções ligam vários tipos de células, como os fibroblastos á matriz extracelular. As porções das integrinas que contactam com a matriz, ligam-se no interior do citoplasma através da sub-unidade Beta aos filamentos de actina que estão unidos a esta sub-unidade por proteínas que ligam a actina a proteínas como a alfa-actinina a talina e a vinculina. |
|
|
COMO E O QUE LIGAM OS HEMIDESMOSSOMAS DA MATRIZ EXTRACELULAR ÁS CÉLULAS? |
Os hemidesmossomas ligam filamentos intermédios através de plectina e integrina á laminina que constitui a lâmina basal. |
|
|
O QUE É A LÂMINA BASAL? |
É o conjunto de moléculas que delimita o tecido epitelial e conjuntivo. |
|
|
QUE FUNÇÃO TÊM AS JUNÇÕES ESTÁVEIS ENTRE CÉLULAS? |
Organizam células em tecidos e facilitam comunicação rápida entre células. |
|
|
PORQUE RAZÃO O TECIDO EPITELIAL QUASE NÃO POSSUI ESPAÇO INTERCELULAR? |
Pois possui muitos tipos de junções entre as células epiteliais que as aproximam e diminuem o espaço entre estas. |
|
|
QUE TIPOS DE JUNÇÕES EXITEM NO TECIDO EPITELIAL? |
Entre as células -Junções de hiato, junções oclusivas, desmossomas, junções de aderência(que formam o cinto de aderência). Entre as células e a lâmina basal - Hemidesmossomas. |
|
|
COMO SÃO E QUE FUNÇÃO TÊM AS JUNÇÕES DE ADERÊNCIA E OS DESMOSSOMAS?QUE PROTEÍNAS INTERVÊM? |
São junções estáveis entre células que envolvem o citoesqueleto. São mediadas pelas selectinas, integrinas, immunoglubulinas e caderinas. |
|
|
COMO É QUE AS CADERINAS INTERVÊEM NAS JUNÇÕES DE ADERÊNCIA? |
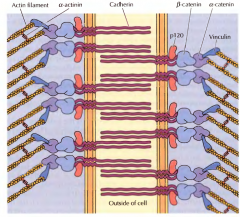
São mediadoras de interações homofilicas( entre moléculas iguais em células diferentes). São responsáveis por manter ligações estáveis entre células em tecidos. São proteínas transmembranares que se ligam-se a beta-catenina que se liga a alfa-catenina que se liga a vinculina que por sua vez se liga a filamentos de actina. |
|
|
QUE ESTRUTURAS FORMAM AS JUNÇÕES DE ADERÊNCIA EM TORNO DAS CÉLULAS EPITELIAIS? |
Um "cinto". |
|
|
COMO É QUE AS CADERINAS INTERVÊEM NOS DESMOSSOMAS? |
As caderinas dos desmossomas chamam-se desmoglina e desmocolina. Ligam-se através de placoglobina e palcofilina aos filamentos intermédios dentro do citosol.
|
|
|
QUE FUNÇÃO TÊM AS JUNÇÕES OCLUSIVAS? |
São junções estáveis e não envolvem elementos do citoesqueleto. Selam o espaço intercelular, impedindo a passagem de moléculas entre entre células. No tecido epitelial intestinal separam o dominio apical(microvilosidades) do domínio basolateral, impedindo a difusão de lípidos e outras moléculas entre estes. Impedem também o movimento de proteínas transmembranares entre os domínios basolateral e apical. |
|
|
PORQUE RAZÃO ESTÃO NORMALMENTE AS JUNÇÕES OCLUSIVAS ASSOCIADAS A JUNÇÕES DE ADERÊNCIA E A DESMOSSOMAS? |
Pois como não estão associadas a elementos do citoesqueleto não têm muita força de aderência. |
|
|
COMO SE FORMAM JUNÇÕES OCLUSIVAS? |
São junções entre proteínas transmembranares do lado externo da membrana que envolvem as células. São formadas por occludina, claudina e JAMs. |
|
|
QUE FUNÇÃO TÊM AS JUNÇÕES DE HIATO? |
Criam canais que permitem a passagem de moléculas entre células mas não de proteínas e ácidos nucleicos. São importantes para sincronizar respostas tecidulares sinalizando outras células do mesmo tecido de uma alteração química (como a presença de iões cálcio) |
|
|
COMO SÃO CONSTITUIDAS AS JUNÇÕES DE HIATO? |
São constituidas por seis conexinas(conexão) que formam um canal entre células. As duas células que comunicam possuem um espaço entre elas.(gap)
|
|
|
EM QUE COMUNICAÇÃO INTERCELULAR SÃO IMPORTANTES AS JUNÇÕES DE HIATO? |
Nas células nervosas, para formar um sinapse. |
|
|
QUE DOENÇA PODE RESULTAR DO MAU FUNCIONAMENTO DAS JUNÇÕES DE HIATO NAS CÉLULAS DO OLHO? |
As cataratas, formam-se precipitados. |
|
|
QUE PROTEÍNAS ENVOLVEM AS INTERAÇÕES TEMPORÁRIAS ENTRE CÉLULAS SEM ENVOLVÊNCIA DO CITOESQUELETO? |
Selectinas, integrinas e hemoglobinas. |
|
|
QUE INTERAÇÕES TEMPORÁRIAS MEDIAM AS SELECTINAS, INTEGRINAS E HEMOGLUBINAS JUNTAS? COMO ACONTECEM? |
Interações temporárias entre leucócitos e plaquetas, a diapedese. As selectinas são ativadas para mover leucócitos para locais de inflamação dos tecidos; na superfície das plaquetas reconhecem glicolípidos dos leucócitos quando estes estão a atravessar a corrente sanguínea. Aderem-se aos leucócitos e depois as integrinas dos leucócitos formam junções mais estáveis ás plaquetas com ajuda de hemoglobinas. Isto permite a diapedese dos leucócitos |
|
|
PORQUE RAZÃO SÃO OS LEUCÓCITOS MAIS LENTOS QUE OS GLÓBULOS VERMELHOS? |
Pois vão formando ligações temporárias ao longo da corrente sanguínea que reduzem a sua velocidade. |
|
|
COMO É FEITA A ADESÃO ENTRE CÉLULAS NOS VEGETAIS? |
Através de uma zona rica em pectinas, a lamela média. |
|
|
QUAL É O EQUIVALENTE ÁS JUNÇÕES DE HIATO NAS CÉLULAS VEGETAIS?
|
Plasmodesmos. |
|
|
COMO SE FORMAM, QUE FUNÇÃO TÊM E COMO SÃO CONSTITUIDOS OS PLASMODESMOS? |
Formam-se por separação incompleta de células durante a mitose. Formam canais em que há continuamento da membrana por onde passa o desmotubulo, que é um prolongamento do REL. |
|
|
QUE MEMBRANAS POSSUEM OS CLOROPLASTOS? |
São constituídos por três sistemas de membranas: as membranas interior e exterior do invólucro cloroplastidial e uma membrana interna a membrana tilacoidal. No interior da membrana tilacoidal existem. |
|
|
COMO É A ORGANIZAÇÃO NO INTERIOR DA MEMBRANA TILACOIDAL? |
A membrana forma discos achatados, chamados de tilacoides, que estão empilhados em montes - grana. Entre os tilacoides existe lumen tilacoidal. |
|
|
O QUE EXISTE NO INTERIOR DO CLOROPLASTO, FORA DA MEMBRANA TILACOIDAL? |
Entre as duas membranas exteriores e inetriores existe espaço intermembranar, o estroma dentro da membrana interior mas fora da membrana tilacoidal |
|
|
COMO SÃO AS CONSTITUIÇÕES E FUNÇÕES DAS MEMBRANAS EXTERIORES E INTERIORES DO CLOROPLASTO? |
A membrana exterior contém porinas e é extremamente permeavel. A membrana interior é extremamente impermeavel mas possui proteínas transportadoras de fosfato e de sacarose. |
|
|
O QUE É E QUE FUNÇÃO TEM O ESTROMA DOS CLOROPLASTOS? |
Consiste no sistema genético do cloroplasto e contém várias enzimas metabólicas, algumas das quais intervêm na fotossíntese. |
|
|
QUAL É A FUNÇÃO DA MEMBRANA TILACOIDAL NOS CLOROPLASTOS? |
A membrana transporta protões para o seu interior, onde existem sistemas de transporte de eletrões. O gradiente eletroquímico formado resulta na síntese de ATP para o estroma. |
|
|
PORQUE RAZÃO É A MEMBRANA TILACOIDAL TÃO FLUIDA? |
Pois possui uma elevada percentagem de glicolípidos insaturados(com ligações duplas) |
|
|
EM QUE DIFEREM OS OUTROS PLASTOS DOS CLOROPLASTOS? O QUE TÊM EM COMUM? |
Contéem todos o mesmo genoma, mas diferem na estrutura e função. Os cloroplastos são os únicos que contém tilacoides. Todos possuem o sistema de duas membranas. |
|
|
COMO SÃO CLASSIFICADO OS DIFERENTES PLASTOS? QUE PLASTOS EXISTEM? |
São classificados quanto ao pigmento. Os cloroplastos contém clorofila, os cromoplastos carotenoides ou xantofilas, os leucoplastos não contém pigmentos e armazenam energia. Os amiloplastos acumulam amido. |
|
|
DE QUE ORGANELOS INDIFERANCIADOS PRESENTES EM REBENTOS E RAIZES DE PLANTAS PROVÉM TODOS OS PLASTOS? |
De proplastos. |
|
|
COMO SE FORMAM OS CROMOPLASTOS? |
A partir de cloroplastos, a clorofila e os tilacoides desintegram-se e são sintetizados carotenoides. |
|
|
QUE TIPO DE PLASTO SE FORMA NA FASE INTERMEDIÁRIA ENTRE OS PROPLASTOS E OS CLOROPLASTOS E QUE SE MANTÉM DESSA FORMA SE NÃO RECEBER LUZ? |
Os Etioplastos. |
|
|
COMO SÃO CONSTITUIDOS OS ETILOPLASTOS? |
Contém apenas protoclorofilida, e um corpo prolamelar. |
|
|
O QUE SE SUCEDE SE UM ETILOPLASTO FOR EXPOSTO A LUZ? |
Os etiloplastos passam a cloroplastos. A protoclorofilida passa a clorofilida e depois com adição de fitol a clorofila. O corpo lamelar passa ao sistema de tilacoides. |
|
|
COMO SE MULTIPLICAM OS PLASTOS? |
Por bipartição. Na zona média dos plastos, acumulam-se proteínas filamentosas que promovem a divisão dos plastos em dois. |
|
|
QUE PERCENTAGEM DAS PROTEÍNAS QUE EXISTEM NO INTERIOR DOS CLOROPLASTOS SÃO SINTETIZADAS POR ESTES? DONDE VÊM AS RESTANTES? |
5-10%, as restantes são sintetizadas pela célula e importadas. |
|
|
COMO SÃO AS PROTEÍNAS IMPORTADAS PARA OS CLOROPLASTOS? |
Um complexo condutor reconhece uma sequência(péptido-trânsito), que consite em 30-100 aminoácidos. Este complexo condutor direcciona a proteína para a o complexo Toc(translocase of the outer cloroplast membrane). No complexo Toc a proteína liga-se a dois recetore membranares. Durante a passagem da proteína pelo complexo Toc, chaperonas Hsp70 mantém a proteína numa estrutura linear. A translocação da proteína também é promovida por fosfatases que estão fixadas no complexo Toc, que hidrolizam o ATP que lhes fornece a energia necessária para translocarem as proteínas. Depois das proteínas passarem o complexo Toc, são transferidas para o complexo da membrana interna o Tic. No lado interno da membrana interna uma chaperona puxa a proteína enquanto esta passa pelo complexo Tic. Quando a proteína atinge o estroma é clivado o péptido-trânsito por uma peptidase processadora do estroma SPP. No estroma, chaperoninas ajudam a proteína a adquirir a sua estrutura terciária. |
|
|
COMO É QUE AS PROTEÍNAS SÃO IMPORTADAS PARA OS TILACOIDES DOS CLOROLASTOS? |
Após a clivagem do péptido-trânsito pela SPP, surge uma segunda sequência sinal que faz a proteína ser dirigida para os tilacoides. Existem três maneiras da proteína ser levada para os tilacoides. A maneira Sec em que a sequência sinal é reconhecida por uma proteína SecA e a proteína com estrutura linear é translocada com energia fornecida pela hidrolização do ATP, através do complexo de translocação Sec. Na segunda maneira(TAT) a proteína é translocada na sua estrutura terciária e depende do gradiente de protões para ser translocada através do complexo TAT. Na terceira forma, as proteínas são reconhecidas por uma chaperona Srp e inserida na membrana do tilacoide. |
|
|
QUAL É O OBJETIVO DA FOTOSSíNTESE?
|
Converter CO2 e H20 em glucose. |
|
|
COMO SE DIVIDE A FOTOSSíNTESE EM DUAS FASES? |
Uma fase dependente da luz em que há síntese de ATP e NADPH e a formação de O2 e H2O, ocorre na membrana tilacoidal. Fase que não depende da luz são usados o ATP e NADPH produzidos na fase dependente da luz para produzir glucose - ocorre no estroma. |
|
|
NA EQUAÇÃO GERAL DA FOTOSSÍNTESE, QUANTAS MOLÉCULAS DE CO2 E H2O ORIGINAM QUANTAS MOLÉCULAS DE GLUCOSE(C6H1206) H20 E O2? |
6 moléculas de CO2 e 12 moléculas de H20 originam uma molécula de glucose(C6H12O6), 6 moléculas de H2O e 6 moléculas de O2. |
|
|
QUAIS SÃO OS PIGMENTOS FOTOSSINTÉTICOS QUE EXISTEM NOS FOTOSSISTEMAS? |
Clorofilas e carotenoides(beta-caroteno) |
|
|
QUAL É A FUNÇÃO DOS PIGMENTOS CAROTENOIDES NOS FOTOSSISTEMAS? |
Protegem as moléculas de clorofila devido á sua estrutura altamente insaturada(poucas ligações duplas) |
|
|
QUAL É O CAMINHO QUE OS FOTÕES FAZEM ATÉ CHEGAR AO CENTRO DE REAÇÃO FOTOSSINTÉTICO? |
Os pigmentos absorvem os fotões e estes são transferidos para o centro de reação com clorofila para fiofitina, depois para quinonas no centro de reação fotossintético |
|
|
QUE TIPOS DE FOSFORILAÇÃO SE SUCEDE NOS TILACOIDES?
|
Fosforilação acíclica e cíclica. |
|
|
QUANTOS FOTOSSISTEMAS EXISTEM NA MEMBRANA DO TILACOIDE?
|
Dois 2 |
|
|
O QUE CONTÉM OS FOTOSSISTEMAS DA MEMBRANA DO TILACOIDE? |
Os pigmentos que absorvem os fotões - a clorofila. |
|
|
O QUE SE SUCEDE NA FOSFORILAÇÃO ACÍCLICA DA FASE LUMINOSA DA FOTOSSÍNTESE? |
Os pigmentos antena absorvem fotões que transferem para os pigmentos de clorofila(centro de reação) e exitam eletrões provenientes da água para niveis de energia superiores. Assim a água é oxidada a oxigénio que é libertado e a protões(perde eletrões). Os eletrões provenientes da água são depois elevados a niveis de energia superiores pelos fotões. Os protões fornecidos pela água acumulam-se no interior do tilacoide.A clorofila depois transfere os eletrões para uma molécula aceitadora de eletrões (Plastoquinona). Da plastpquinona os eletrões são transferidos para o Citocromo bf que com a energia dos eletrões bombeia protões para o interior do tilacoide. Depois os eletrões seguem para a plastocianina e da plastocianina para o fotossistema I. No fotossistema I os eletrões são de novo exitados para niveis de energia superiores e são transferidos para a ferrodoxina(proteína) que complexada com a enzima NADP reductase reduz o NADP a NADPH que vai ser utilizado no ciclo de calvin. O gradiente de protões que é criado no interior do tilacoide faz com que estes depois passem por a ATP sintase que produz energia(ATP) para o ciclo de calvin(fase escura). |
|
|
QUE NOME SE DÁ AO CONJUNTO DE PROTEÍNAS NA MEMBRANA DO TILACOIDE? |
Cadeia transportadora de eletrões. |
|
|
O QUE SE SUCEDE DURANTE A FOSFORILAÇÃO CÍCLICA DA FASE LUMINOSA DA FOTOSSÍNTESE NOS TILACOIDES? |
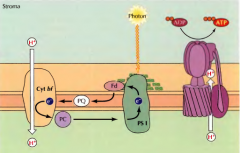
No final da cadeia transportadora de eletrões no fotossistema I não é reduzido NADP e os eletrões passam para o citocromo bf e para o fotossistema II sendo bombeados mais protões o que promove um maior gradiente de protões no interior do tilacoide e uma maior síntese de ATP por parte da célula. |
|
|
COMO ESTÁ ORGANIZADA A MEMBRANA TILACOIDAL?
|
É uma membrana muito comprida pela qual estão espalhaods complexos envolvidos na fosforilação. |
|
|
O QUE OCORRE DURANTE O CICLO DE CALVIN? |
Numa primeira fase fixa-se o dixódio de carbono e uma enzima, a RuBisco, junta o dióxido de carbono á RuBP(ribulose bi-fosfato)(com 5 carbonos), ficando 6 moléculas de 6 carbonos(fosfoglicerado). Na segunda fase, da redução que se divide em duas moléculas de 3 carbonos com o uso de 6 ATP e com a oxidação de 6 NADPH a NADP fornecendo o H á molécula , formam-se 6 moléculas de gliceraldeído tri-fosfato(G3P). Uma das G3P é depois usada para produzir glucose. As outras 5 G3P são reutilizadas com uso de 3ATP para a regeneração da RuBP. |
|
|
QUE IMPORTANTE MECANISMO BIOLÓGICO DESEMPENHA A MITOCÔNDRIA QUE PRODUZ ENERGIA NA FORMA DE ATP PARA A CÉLULA? |
Fosforilação oxidativa |
|
|
COMO É CONSTITUIDA A MITOCÔNDRIA? |
Membrana externa, espaço intermebranar, mebrana interna. A membrana interna forma dobras - cristas mitocôndriais. No interior da mitocôndria existe matriz. Na matriz estão presentes o material genético mitocondrial e enzimas importantes para a fosforilação oxidativa. |
|
|
COMO SÃO CONSTITUIDAS A MEMBRANA EXTERNA E INTERNA DA MITOCÔNDRIA?
|
A membrana externa é constituída por porinas e a membrana interna constituida por muitas proteínas importantes para a transferência de metabolitos celulares. |
|
|
COMO SE DIVIDEM AS MITOCÔNDRIAS? |
Por bipartição |
|
|
QUE UTILIDADE TÊM AS CRISTAS MITOCONDRIAIS? |
Aumenta a superfície interna da mitocôndria. |
|
|
COMO É O GENOMA MITOCONDRIAL? |
O genoma mitocondrial codifica apenas 5-10% das proteínas mitocondriais. È constituido por DNA circular, em que a maior parte das sequências não codifica proteínas. Os codões do genoma mitocondrial humano podem ter significados diferentes do genoma humano apesar de terem a mesma sequência de aminoácidos. |
|
|
COMO É QUE AS PROTEÍNAS DO CITOSOL SÃO IMPORTADAS PARA A MITOCÔNDRIA?
|
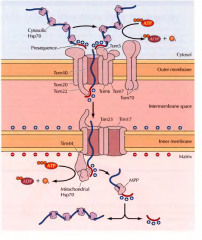
As proteínas que vão ser importadas possuem uma pré-sequência de aminoácidos que é reconhecidas por chaperonas que as encaminha até a membrana. Na parte exterior da membrana, recetores membranares reconhecem a proteína. Estes recetores pertencem aos complexos transmembranares Tom. A proteína é assim translocada para o espaço intermembranar mitocondrial através dos complexos Tom com uso de ATP. Depois as proteínas são translocadas através da membrana interna pelo complexo de translocação Tim. Esta segunda translocação é possivel devido á diferença de potencial entre o espaço intermebranar(positivo) e o espaço interior da mitocôndria(negativo). Esta diferença de potencial deve-se á transferência de protões resultantes da fosforilação oxidativa para o espaço intermembranar. Ao serem translocadas pela membrana interna as proteínas estão já com a sua estrutura linear(secundária). São as chaperonas que mantêm as proteínas lineares até á membrana externa. Uma chaperona na parte interna da membrana interna ajuda na importação das proteínas com energia fornecida pelo ATP. No interior da mitocôndria uma peptidase MPP cliva a pré-sequência e chaperonas e chaperoninas ajudam a proteína a adquirir a sua estrutura terciária. |
|
|
COMO É QUE AS PROTEÍNAS DA MEMBRANA INTERNA DA MITOCÔNDRIA SÃO INSERIDAS? |
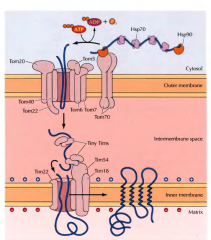
As proteínas com este destino não possuem pré-sequências, possuem multiplas sequências internas de importação que são reconhecidos pelas chaperonas que são reconhecidos por recetores da membrana externa das mitocondrias e permitem a sua passagem para o espaço intermembranar. No espaço intermembranar pequenas proteínas(pequenos-Tim) reconhecem a proteína e levam-na para o complexo Tim para ser parcialmente translocada até os seus sinais-stop provocarem a paragem na translocação e a inserção na membrana . |
|
|
DE QUE FORMAS PROVÊM AS PROTEÍNAS QUE VÃO SER INSERIDAS NAS MEMBRANAS? |
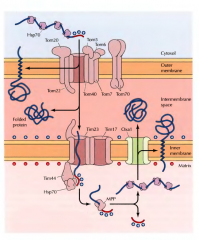
Algumas possuem pré-sequências e sequências de sinal internas, sendo assimtranslocadas para o espaço intermembranar. Algumas proteínas são inseridas na membrana externa. Outras proteínas permanecem no espaço intracelular. Algumas proteínas são translocadas para o interior da mitocôndria, mas quando clivadas revela-se um péptido sinal que as leva a voltar para serem inseridas numa das membranas através do complexo de translocação Oxa1 ou para serem levadas para o espaço intermebranar. Também é através do complexo de translocação Oxa1 que as proteínas que são sintetizadas no interior da mitocôndria são translocadas . |
|
|
DE QUE FORMAS PROVÊM AS PROTEÍNAS QUE VÃO SER INSERIDAS NAS MEMBRANAS DA MITOCÔNDRIA? |
Algumas possuem pré-sequências e sequências de sinal internas, sendo assimtranslocadas para o espaço intermembranar. Algumas proteínas são inseridas na membrana externa. Outras proteínas permanecem no espaço intracelular. Algumas proteínas são translocadas para o interior da mitocôndria, mas quando clivadas revela-se um péptido sinal que as leva a voltar para serem inseridas numa das membranas através do complexo de translocação Oxa1 ou para serem levadas para o espaço intermebranar. Também é através do complexo de translocação Oxa1 que as proteínas que são sintetizadas no interior da mitocôndria são translocadas . |
|
|
QUAL É O FOSFOLÍPIDO MAIS IMPORTANTE DA MITOCÔNDRIA? COMO É OBTIDO? |
É a cardiolipina, que contém quatro cadeias de ácidos gordos. É sintetizada após a transferência de fosfatidilcolina e fosfatildiletanolamina para a mitocôndria. É responsável pela resitência á oxidação da membrana da mitocôndria. |
|
|
O QUE OCORRE ANTES DA FOSFORILAÇÃO OXIDATIVA NAS MITOCÔNDRIAS? |
Primeiro ocorre a glicólise no citosol e a glicose é convertida em piruvato. O piruvato(ácido pirúvico) é atravessa depois a membrana através de porinas e de proteínas membranares e chega á matriz mitocondrial. Na matriz da mitocôndria o ácido pirùvico é oxidado acetil CoA(coenzima-A) que é desdobrada em CO2 através de enzimas no ciclo de ácido cítrico. A acetil CoA também é obtida através da oxidação de ácidos gordos no ciclo de ácido cítrico(ciclo de krebs). Ao mesmo tempo dá-se a redução de NAD e FAD a NADH eFADH2. De seguida dá-se a forforilação oxidativa. |
|
|
COMO OCORRE A FOSFORILAÇÃO OXIDATIVA NAS MITOCÔNDRIAS? |
Durante a fosforilação oxidativa os eletrões do NADH e FADH2 combinam-se com o O2 e as reações de redução e oxidação que ocorrem resultam na síntese de ATP. Os eletrões do NADH entram para a cadeia transportadora de eletrões através do complexo I( NADH desidrogenase), os eletrões são transferidos para o mononucleotideo de flavina(FMN) e depois através de um complexo de Ferro-Enxofre(F-S) para a coenzima Q(Ubiquinona). Esta última é uma proteína que transfere eletrões para o complexo III(citocromo bc1). Os eletrões passam depois do citocromo bc1 para o citocromo c e depois para o citocromo c oxidase(complexo IV) onde são transferidos para o oxigénio e é produzida água. Outro complexo(II), o Succinato desidrogenase, recebe eletrões do FADH2, que é convertido em FAD que os transfere para outra coenzima independente Q que os transfere para o o citocromo bc1 e fazem o seu percurso normal. Durante este processo acumulam-se protões no espaço intermebranar que entram na ATP sintase do para o interior da membrana e promovem a sintese de ATP. |
|
|
DEPENDENDO DAS CONDIÇÕES AMBIENTAIS, QUE OUTRA FUNÇÃO PODEM TER AS ATP-ASES PARA ALÉM DA PRODUÇÃO DE ATP?
|
Podem assumir a função de bombas tipo F e bombear protões contra o gradiente de concentração produzindo ADP apartir de ATP |
|
|
QUE DOENÇA É PROVOCADA POR UMA MUTAÇÃO NO DNA MITOCONDRIAL QUE PROVOCA CEGUEIRA? COMO OCORRE? |
È a neuropatia óptica hereditária de Leber. A mutação leva a alterações na NADH desidrogenase que provoca o mau funcionamento da fosforilação oxidativa e produção de ATP isuficiente. |
|
|
COMO É QUE A MITOCÔNDRIA PROVOCA O STRESS OXIDATIVO? |
A oxidação excessiva da cardiolipina leva á libertação do citocromo C. Formam-se poros na mitocôndria e transfere-se o citocromo c para o exterior do organelo, liga-se á proteína Apaf-1 formam-se apoptossomas e são ativadas caspases que provocam a morte celular. |
|
|
O QUE SÃO OS PEROXISSOMAS? |
São organelos com uma única membrana, que contêm enzimas envolvidas em varios processos metabólicos. Não possuem o seu próprio material genético e todas as suas proteínas(peroxinas) são importadas. Podem-se multiplicar por bipartição e podem-se regenerar. Podem possuir estruturas cristalinas no seu interior. |
|
|
COMO SE FORMAM OS PEROXISSOMAS? |
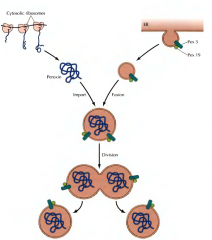
Tudo começa no RE, onde são formadas proteínas percursoras. Estas proteínas interagem e formam vesículas que se separam do RE e se fundem umas com as outras ou com peroxissomas já pré-existentes. São depois importadas proteínas e lípidos e forma-se um peroxissoma maturo. |
|
|
COMO SÃO IMPORTADAS AS PROTEÍNAS PARA OS PEROXISSOMAS? |
Existem duas formas. As proteínas possuem uma sequência de aminoácidos Ser-Lys-Leu (ou outras) que é reconhecida por chaperonas que as guiam para recetores membranares dos peroxissomas que reconhecendo as proteínas permitem a sua passagem. As proteínas passam assim por um complexo de translocação na sua estrutura secundária e no interior atigem a estrutura terciária. A suas sequências de sinal não são clivadas. |
|
|
QUE FUNÇÕES TÊM OS PEROXISSOMAS? |
Oxidam ácidos gordos, síntetizam colesterol e dolicol, síntetizam ácidos biliares no fígado e plasmalogénios, catabolizam purinas e convertem aminoácidos em hidratos de carbono. |
|
|
COMO É QUE OS PEROXISSOMAS COMPLEMENTAM AS MITOCÔNDRIAS? |
Oxidam ácidos gordos que vão ser utilizados pela mitocôndria. |
|
|
QUE IMPORTÂNCIA TEM A ENZIMA CATALASE NOS PEROXISSOMAS? |
Decompões o peróxido de hidrogénio convertendo-o em água ou utilizando-o para oxidar outros compostos. |
|
|
DÁ UM EXEMPLO DE COMO OS PEROXISSOMAS OXIDAM ÁCIDOS GORDOS? |
Oxida a acetil CoA produzindo peróxido de hidrogénio que depois irá ser utilizado para oxidar outro composto orgânico. |
|
|
EXPLICA UMA RELAÇÃO ENTRE OS PEROXISSOMAS E AS MITOCÔNDRIAS |
Os ácido gordos entram no peroxissoma e são oxidados a Succianato que depois é utlizdo na mitocôndria, durante o ciclo de Krebs. |
|
|
QUE IMPORTANTE FUNÇÃO TÊM OS PEROXISSOMAS NO FÍGADO? |
Síntetizam o colesterol. |
|
|
QUE IMPORTANTES REAÇÕES DESEMPENHAM OS PEROXISSOMAS NAS CÉLULAS VEGETAIS? |
Intervêm no ciclo de glioxilato que converte ácidos gordos em hidratos de carbono- glicolato (que vai ser usado pelos cloroplastos) e glicina( a ser usada pela mitocôndria e convertida em serina. Os peroxissomas també desempenham funções importantes na fotorespiração: Por vezes no cloroplasto é utilizado O2 em vez de CO2 no ciclo de calvin produzindo uma molécula de fosfoglicolato, que não é utilizada pelo cloroplasto e é convertida em glicolato que é transferido para o peroxissoma onde é oxidado e convertido em glicina. A glicina é transferida para a mitocôndria onde é convertida a serina libertando CO2 e NH3(que é transferido para fora da mitocôndria através das transamninases). A serina depois volta para o peroxissoma e é convertida a glicerato que irá ser utilizado no ciclo de Calvin nos cloroplastos. |
|
|
QUE DOENÇA CONHECES QUE É CAUSADA POR ANOMALIAS NOS PEROXISSOMAS? |
Síndroma de Zellweger - Existem poucos ou nenhuns peroxissomas, o que resulta me vários problemas no oragnismo. |
|
|
QUE TIPOS DE SINALIZAÇÃO CELULAR EXISTEM? |
Sinalização celular entre direta entre células, sinalização celular através da secreção de moléculas(sistema endócrino, parácrino e autócrino). |
|
|
QUE TIPOS DE SINALIZAÇÃO DIRETA ENTRE CÉLULAS CONHECES? |
As integrinas e caderinas ligam-se a células e á matriz-extracelular regulando as interações e proliferação celular. |
|
|
EM QUE CONSISTE A SINALIZAÇÃO CELULAR ENDOCRINA?
|
Secreção de hormonas para a corrente sanguínea atuando longe de onde são libertadas |
|
|
EM QUE CONSISTE A SINALIZAÇÃO CELULAR PARACRINA? |
Uma molécula libertada por uma célula atua numa célula próxima desta. É o caso dos neurotransmissores libertados entre células nervosas através de sinapse. |
|
|
EM QUE CONSISTE A SINALIZAÇÃO CELULAR AUTÓCRINA? |
É a resposta de células a moléculas que elas próprias produzem. Como é o caso dos linfócitos-T que produzem moléculas que causam um aumento na sua taxa de divisão. |
|
|
O QUE TORNA A SINALIZAÇÃO ATRAVÉS DE HORMONAS ESTERÓIDES DIFERENTE DOS OUTROS TIPOS DE SINALIZAÇÃO? |
Algumas células possuem recteptores de sinalização no seu interior sendo apenas ativados por moléculas que são capazes de se difundir na célula como é o caso das Hormonas Esteroides. |
|
|
QUE TIPO DE ESTEROIDES CONHECES? COMO SÃO PRODUZIDOS? ONDE SÃO SINTETIZADOS?QUE OUTRAS HORMONAS FUNCIONAM DE FORMA SEMELHANTE AOS ESTEROIDES? |
Todos os esteroides são sintetizados a partir de colesterol. A testoesterona o estrogénio e a progesterona são produzidos pelas gónadas. Os corticoesteroides(cortisol e aldoesterona) pelas glândulas adrenais e atuam nos rins regulando os niveis de água e sal. A hormona da tiroide a vitamina D3 e o ácido retinoico não são esteroides mas desempenham funções semelhantes. |
|
|
COMO ENTRAM AS HORMONAS ESTEROIDES, DA TIROIDE, VITAMINA D3 E ÁCIDO RETINOICO PARA O INTERIOR DA MEMBRANA? ONDE SE LIGAM NO INTERIOR? |
Devido ao seu carater hidrofobico diffundem-se através da membrana plasmática. Ligam-se a recetores nucleares. |
|
|
QUE AÇÃO TEM O ESTROGÉNIO A NIVEL DE SINALIZAÇÃO CELULAR? COMO SE PROCESSA? O QUE SE SUCEDE NA SUA AUSÊNCIA? |
O estrogénio difunde-se através da membrana plasmática, liga-se ao recetor de estrogénio(que estava associado a um chaperona que se separa após a ligação), que se liga ao DNA. Uma molécula HAT(histona acetil transferase) funciona como coativador e ativa a transcrição de DNA. Na ausência de estrogénio o recetore associa-se na mesma ao DNA mas um corepressor fica também associdado(HDAC) e impede a transcrição. |
|
|
COMO FUNCIONA A SINALIZAÇÃO CELULAR ATRAVÉS DO ÓXIDO NÍTRICO? |
Consiste em sinalização paracrina. O NO(óxido nítrico) difunde-se através da membrana das células(é um gás). No interior altera a atividade de enzimas. |
|
|
COMO É SINTETIZADO O ÓXIDO NÍTRICO QUE VAI AGIR NA SINALIZAÇÃO PARÁCRINA? |
É sintetizado a partir do amino acido arginina atarvés da enzima óxido nítrico sintaze. |
|
|
COMO É QUE O ÓXIDO NÍTRICO REGULA A VASODILATAÇÃO? |
Primeiro libertam-se neurotransmissores que atuam em células especificas e estimulam a sintese de NO. Este difunde-se com células musculares e ativa o mecanismo de vasodilatação. |
|
|
COMO SE DÁ A SINALIZAÇÃO CELULAR ATRAVÉS DE NEUROTRANSMISSORES? |
Um potencial de ação atinge o terminal do neurónio que provova a libertação de neurotransmissores através do sinapse. Sendo moléculas hidrofilicas os neurotransmissores não atravessam a membrana e ligam-se a recetores membranares resultando na abertura de canais de iões. |
|
|
O QUE SÃO FATORES DE CRESCIMENTO? |
São moléculas sinalizadoras que controlam o crescimento e diferenciação celular. |
|
|
COMO FUNCIONA A AÇÃO DOS FATORES DE CRESCIMENTO DAS PLAQUETAS(PDGF)? |
O PDGF é aramzenado em plaquetas de sangue e libertado durante a coagulação de sangue no local da ferida que estimula a proliferação de fibroblastos junto á ferida e promove o crescimento do tecido. |
|
|
COMO É REGULADO O DESENVOLVIMENTO E CRESCIMENTO DAS PLANTAS? |
Através de hormonas |
|
|
QUE TIPOS DE HORMONAS EXISTEM NAS PLANTAS? |
Auxinas, gibberelinasm citoquininas, acido absíssico, e etileno. |
|
|
QUE EFEITO TEM O ÁCIDO INDOLE-ACÉTICO(AUXINA) NAS PLANTAS? |
Promove o crescimento das células ao remover as ligações das hemiceluloses à celulose. A auxina também se liga a um recetor chamado de Ubiquitina ligase que causa a degradação de proteínas repressoras de genes e provoca a transcrição de genes específicos. |
|
|
COMO É CONSTITUDO O COMPLEXO PROTEÍNA G-RECEPTOR ? |
Por sete helices alfa transmenbranares que estão associadas a um recetor membranar. |
|
|
COMO OCORRE A ATIVAÇÃO E FUNCIONAMENTO DE UMA PROTEÍNA G? |
As moléculas(ligandos) ligam-se aos recetores da parte extracelular e causam alterações na proteína que permitem que a parte citosólica(no interior da membrana) do recetor se ligue á proteína G. Isto ativa a proteína G que se dissocia do recetor e transfere o sinal para o seu destino(enzima ou canal de iões). |
|
|
COMO É REGULADA A SÍNTESE DE cAMP(AMP CÍCLICO)? |
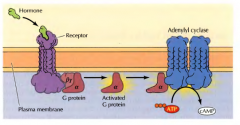
Após a ativação da proteína G, esta ativa uma enzima(adenilil ciclase que transforma ATP em cAMP) |
|
|
COMO É FEITA A REGULAÇÃO DAS PROTEÍNAS G NA LIBERTAÇÃO DA HORMONA EPINEFRINA? |
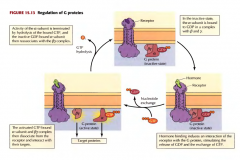
As proteínas G são constituidas por três sub-unidades: alfa, beta e gama. Na fase de descanso a unidade alfa está a fixar o GDP e forma um complexo com as sub-unidades beta e gama. Quando uma hormona se liga ao recetor, este interage com a proteína desencadeando a troca de GDP na unidade alfa por GTP. A subunidade alfa depois separa-se das outras subunidades. Assim tanto a sub-unidade alfa com GTP como o complexo beta-gama interagem com a célula desencadeando uma resposta celular. No final o GTP na sub-unidade alfa é hidrolizado a GDP e a sub-unidade volta a formar um complexo com as sub-unidades beta e gama. |
|
|
EM QUE ESTÃO ENVOLVIDOS OS RECEPTORES DE CINASES DA TIROSINA? |
Na ativação de enzimas intracelulares, crescimento celular e diferenciação. |
|
|
COMO SÃO COMPOSTOS OS RECEPTORES CINASES DA TIROSINA? |
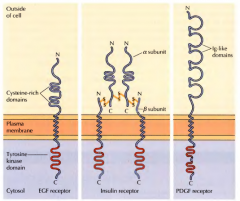
Uma porção citosólica do recetor com cistenina. Uma parte inserida na membrana e um domínio cinase tirosina no interior da célula. Alguns recetores consistem em duas cadeias de polipeptidos e outros possuem um dominio de imunoglubina no exterior da célula e dois dominios de tirosina cinase no interior da célula. |
|
|
COMO OCORRE A SINALIZAÇÃO ATRAVÉS DOS RECETORES CINASES DA TIROSINA NO CASO DA INSULINA? |
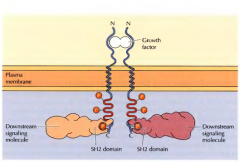
Um factor de crescimento liga-se a ambas as cadeias polipeptídicas o que provoca a dimerização. Depois ocorre a autofosforilação e as duas cadeias polipetídicas fazem a fosforilação cruzada da tirosina nas porções intracelulares dos recetores. Esta autofosforilação aumenta a atividade a proteína cinase e cria locais de ligação para outras proteínas transmitirem sinais intracelulares. As proteínas que se ligam a estes locais têm o nome de SH2. |
|
|
COMO SE FORMA O cAMP? |
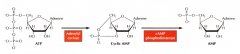
Forma-se a partir do ATP através da adenilil ciclase e é degradado a AMP através da cAMP fosfodiesterase. |
|
|
COMO FUNCIONA A VIA DE TRANSDUÇÃO DO SINAL INTRACELULAR NO CASO DO cAMP? |
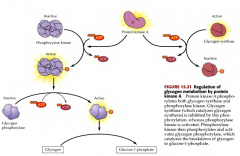
Depois do cAMP ter sido covertido atarvés do ATP. O cAMP funciona como segundo mensageiro no sinalização hormonal da epinefrina. O cAMP liga-se a uma enzima (proteína cinase A)(que é composta por quatro sub-unidades) que leva á dissociação de duas sub-unidades que iram fosforilar resíduos de serina na proteínas em que agirem. A proteína cinase A fosforiliza a fosforilaze cinase que ficando ativa vai fosforilar a fosforilaze glicogénio que decompõe o glicogénio em glicogénio-1-fosfato. A proteína cinase também fosforiliza a glicogénio sintase que cataliza a sintese de glicogénio |
|
|
DÁ UM EXEMPLO DE UMA AMPLIFICAÇÃO DE SINAL CELULAR |
Uma molécula sinalizadora a epinefrina(hormona) liga-se a recetor ligado a proteína G que provoca uma via de transdução que varias enzimas e proteínas são ativadas, o que faz com que a glucose seja sintetizada a partiir de glicogénio. |
|
|
COMO É CONSTITUIDO O INVÓLUCRO NUCLEAR? |
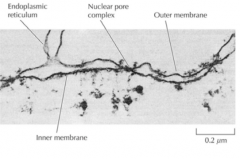
É constituido por duas membranas: interna e externa (que é extensão do RE) e por complexos de poros nucleares. A membrana externa é semelhante á membrana do RE e possui ribossomas na sua superfície. |
|
|
QUE FUNÇÕES TÊM AS MEMBRANAS DO INVÓLUCRO NUCLEAR? COMO SE UNEM? |
Funcionam como barreiras e separam os contéudos do núcleo do citoplasma. As duas membranas têm constituição semelhante a outras membranas celulares e são constituidas por uma bicamada fosfolípidica. As duas membranas unem-se nos complexos de poros nucleares, através duas quais macromoléculas entram no núcleo. |
|
|
NUMA VISUALIZAÇÃO DO NÚCLEO ONDE SE PODEM ENCONTRAR OS COMPLEXOS DE POROS NUCLEARES? |
Na zona em que a cromatina está menos condensada: junto á Eucromatina. |
|
|
O QUE ESTÁ JUNTO Á MEMBRANA INTERNA DO NÚCLEO? |
A lâmina nuclear. |
|
|
COMO É CONSTITUIDA A LÂMINA NUCLEAR? |
Por filamentos intermédios de lamina nucleares. Estas associam-se a proteínas membranares da membrana interna nuclear (emerina e lamina B) que as permitem organizar dentro do núcleo. Também estão ligadas á cromatina. |
|
|
O QUE FOSFORILIZA DURANTE A PROFASE (DESORGANIZAÇÃO NUCLEAR) |
O invólucro nuclear. |
|
|
COMO SÃO CONSTITUIDOS OS COMPLEXOS DE POROS NUCLEARES? |
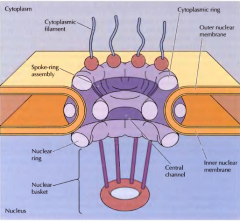
Por filamentos citoplasmáticos, anel citoplasmático, anel central, anel nuclear e cesto nuclear. É constituido por cerca de 50 proteínas(nucleoporinas) |
|
|
QUE FUNÇÃO TÊM OS COMPLEXOS DE POROS NUCLEARES? |
Permitem a transferência de moléculas e DNA para dentro e fora do núcleo. |
|
|
QUE DUAS FORMAS DE TRANSPORTE INTERMENBRANAR EXISTEM NO NÚCLEO? |
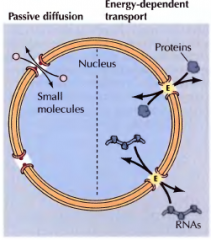
Difusão passiva de pequenas moléculas e transporte com gasto de ATP de proteínas e RNA. |
|
|
QUE SEQUÊNCIAS DE AMINOÁCIDOS POSSUEM AS PROTEÍNAS DESTINADAS AO NÚCLEO? |
Nuclear Localization Signals (NLS). |
|
|
COMO É QUE AS PROTEÍNAS SÃO IMPORTADAS PARA O NÚCLEO? |
As sequências de aminoacidos NLS são reconhecidas por proteínas que funcionam como recetores de transporte nuclear(importinas) que se ligam a proteínas da extremidade do poro nuclear, e de seguida as importinas transportam as proteínas para o interior da membrana. |
|
|
COMO É QUE AS MACROMOLÉCULAS SÃO TRANSPORTADAS PARA O INTERIOR DO NÚCLEO? |
Através da proteína RanGDP. |
|
|
COMO É QUE A RANGTP INTERVEM NA EXPORTAÇÃO DE IMPORTINAS DO NÚCLEO? |
A RanGTP liga-se à importina libertando a proteína que esta importou. A RanGTP depois exporta a importina para o exterior do núcleo e o GTP é hidrodlizado a GDP pela Ran GAP.
|
|
|
COMO É QUE AS PROTEÍNAS SÃO EXPORTADAS DO NÚCLEO? |
Possuem sequência de aminoácidos NES(nuclear export signals) que são reconhecidos por exportinas e estas exportam-nas. O RanGTP liga-se ao complexo da exportina+proteína e estabiliza-o(em contraste com a importação). A hidrólise do RanGTP no citosol faz com que a proteína se solte do complexo e se liberte no citoplasma. |
|
|
COMO É A ESTRUTURA DA CROMATINA NO NÚCLEO INTERFÁSICO? |
Possui zonas muito condensadas(heterocromatina) e zonas pouco condensadas(eucromatina). |
|
|
QUE DOIS TIPOS DE HETEROCROMATINA EXISTEM E QUAL A DIFERENÇA ENTRE ESTES? |
Heterocromatina constitutiva que contém sequências de DNA não são transcritas. Heterocromatina facultativa que pode se descondensar e ser transcrita. |
|
|
COMO É A ESTRUTURA DA CROMATINA? |
O DNA enrola-se a volta das histonas e de proteínas básicas. O DNA + histonas forma nucleossomas que se associam e formam a cromatina. |
|
|
EM RESUMIDO,COMO SE FORMAM PROTEÍNAS PRIMÁRIAS APARTIR DO DNA? |
DNA é transcrito para mRNA(RNA mensageiro) depois é processado por ribossomas para rRNA(RNA ribossomal) que após processamento passa a tRNA que é traduzido originando as proteínas. |
|
|
COMO OCORRE A REPLICAÇÃO DE DNA?(FORQUILHA DE REPLICAÇÃO) |
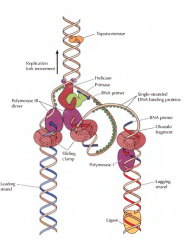
O DNA é replicado sempre na sequência carbono-5 ---- carbono-3(5' 3') . A cadeia de DNA só é replicada de maneira continua numa das cadeias. A helicase separa as duas cadeias uma das quais é sintetizada continuamente. Na outra cadeia: A primase sintetiza pequenos fragmentos de RNA. Estes vão se ligar á cadeia de DNA (com ajuda da DNA polimerase) que foi separada pela helicase e formam fragmentos de Okasaki que são unidos através da DNAligase. Estes fragmentos contêm uma cadeia de RNA e uma cadeia de DNA |
|
|
COMO É CONSTITUIDO UM CROMOSSOMA DURANTE A METAFASE? |
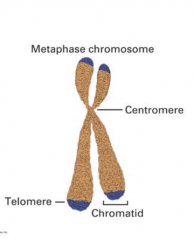
Por dois cromatídeos um cetrómero no meio e telomeros nas pontas. Os cinetocoros são olocal de ligação do fuso acromático aos centrómeros. |
|
|
O QUE SE SITUA NO NUCLÉOLO? |
O rDNA(DNA para a transcrição de proteínas que iram formar os ribossomas), este é o local onde se dá a transcrição de rRNA. |
|
|
O QUE SÃO ORGANIZADORES NUCLEOLARES? |
São regiões de um cromossoma que contêm rDNA. |
|
|
COMO É CONSTITUIDO O NUCLÉOLO? |
Por três regiões: Componente granular(contém sub-unidades ribossomais em diferentes estádios de formação), centros fibrilhares(contêm o rDNA), componente fibrilhar densa que contém o pré-rRNA e proteínas associadas. |
|
|
COMO SE FORMAM OS RIBOSSOMAS? |
Primeiro é necessária a formação de rRNA a partir de proteínas ribossomais e rRNA 5S. Para isto a RNA polimerase II transcreve os genes que codificam as proteínas ribossomais fora do núcleo e a RNA polimerase III transcreve os genes do rRNA 5S. As proteínas ribossomais e o rRNA 5S são depois transportadas para o núcleo onde se associam a moléculas de rRNA para formarem partículas pré-ribossomais. Depois há clivagem do pre-RNA e as partículas pré-ribossomais são exportadas como sub-unidades 40S e 60S. |
|
|
COMO OCORRE A DESORGANIZAÇÃO DO INVÓLUCRO NUCLEAR DURANTE A MITOSE? |
As laminas da lâmina nuclear são fosforiladas pela Cdk1 o que resulta na despolarização da lâmina nuclear durante o final da Profase. O RE absorve o involucro nuclear. No fim da mitose o invólucro volta a formar-se com ajuda das laminas. |