![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
213 Cards in this Set
- Front
- Back
- 3rd side (hint)
|
Anucleate Cell |
Bacteria lacking a nucleoid but have similar shape to wild type |
|
|
|
Apoptosis |
Cellular suicide through signaling pathways |
|
|
|
Checkpoint |
Biochemical control mechanism preventing cell from progressing to next stage of replication unless specific requirements are met |
|
|
|
Cyclins |
Proteins that bind and help activate CDKs. Concentration varies throughout cell cycle, periodic availability regulates cycle progression |
|
|
|
Cyclin-Dependent Kinase |
Kinase family that are inactive unless bound to cyclin. Most participate in cell cycle regulation |
|
|
|
Doubling Time |
Period of time that it takes for bacterial cell to reproduce C + D ~ 60 min E. coli can be 18-180 min |
|
|
|
Minicell |
Anucleate bacterial cell produced by division that generates a cytoplasm without a nucleus |
|
|
|
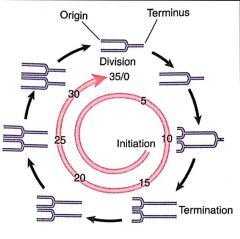
Multiforked Chromosome |
Bacterial chromosome that has more than one set of replication forks, because a second initiation has occurred before the first cycle of replication has been completed |
|
|
|
Nucleoid |
The structure in a prokaryotic cell that contains the genome, DNA is bound to proteins, not enclosed by a membrane |
|
|
|
Oncogene |
A gene, when altered, may cause cancer. Typically dominant mutation. |
|
|
|
Replicon |
Unit of replication One in prokaryote, multiple in eukaryote |
|
|
|
Restriction Point |
The point during G1 at which a cell becomes committed to division. (In yeast this point is known as START). |
|
|
|
S-Phase |
Restricted part of eukaryotic cell cycle where DNA synthesis occurs |
|
|
|
Septal Ring |
Complex of several proteins coded by fts genes of E. coli that forms at cell midpoint. Gives rise to septum at cell division. First protein to be incorporated is ftsZ, where original Z-ring name comes from. |

|
|
|
Septum |
Peptidoglycan structure that forms in center of dividing bacterium, provides site at which daughter bacteria separate. Same term used for wall between plant cells at end of mitosis. |
|
|
|
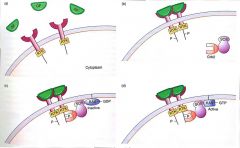
Signal Transduction Pathway |
Process by which a stimulus or cellular state is sensed by and transmitted to pathways within cell |
|
|
|
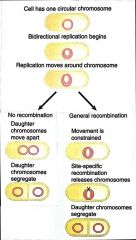
Site-Specific Recombination |
Recombination that occurs between 2 specific sequences |
|
|
|
Topoisomerase |
Enzyme that changes the number of times the 2 strands in a closed DNA molecule cross each other. Cuts DNA, passes DNA through the break, and reseals the DNA |
|
|
|
Tumor Suppresor |
Proteins that usually act by blocking cell proliferation or promoting cell death. Cancer may result when tumor suppressor gene is inactivated or functions improperly Recessive loss of function mutations |
|
|
|
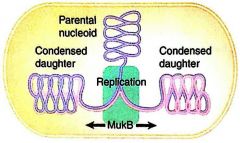
mukA |
Prokaryotes Attaches chromosome to envelope |
|
|
|
mukBEF |
Prokaryotes Condenses the chromosome Protein complex |
|
|
|
ftsZ |
Prokaryotes Involved in septum formation Temperature sensitive Early cell cycle - everywhere in cytoplasm Just before division - at midpoint Hyperactivity causes increased septum formation, minicells, potentially anucleate cells No activity or hypoactivity causes filamentous bacteria without septums Associates with 2 proteins independently: zipA - embedded in membrane on cytosolic side, allows ftsZ to bind to membrane ftsA - cytosolic protein |
|
|
|
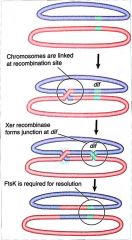
ftsK |
Prokaryotes Xer site-specific recombination. Breaks up Holliday junction, which is at dif site (28 bp). ftsK responsible for site-specific recombination. Cannot occur without it. Located in septum. |
|
|
|
Rod Shape Factors |
Prokaryotes MreB PBP2 (Penicillin binding protein) RodA |
|
|
|
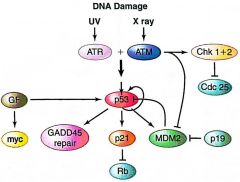
ATM |
DNA damage pathway
Kinase activated by x-ray Activates pathway Acts on Chk1/2 and p53 |
|
|
|
ATR |
DNA damage pathway
Kinase activated by UV Activates pathway Acts on Chk1/2 and p53 |
|
|
|
Chk1 and Chk2 |
DNA damage pathway
Activated by ATR or ATM
Inhibit cell growth Act on/inhibit CDC25 |
|
|
|
p21 |
DNA damage pathway
Inhibits CDK/Cyclin complex
Activated by p53 Acts on cyclins and CDKs |
|
|
|
p53 |
DNA damage pathway
Major tumor suppressor
Activated by ATR or ATM Acts on p21 and MDM2 |
|
|
|
GF |
DNA damage pathway
Growth factor Cell cycle stimulation Activates p19 transcription
Activated by cell cycle progression Acts on MYC |
|
|
|
MDM2 |
DNA damage pathway
Inhibits p53 via negative feedback loop Targets it to ubiquitin-dependent proteosomal degredation pathway Inhibited by DNA damage
Acted on by p53 Acts on p53 |
|
|
|
CDC25 |
DNA damage pathway
Phosphatase that removes inhibitory phosphate groups to activate CDK
Acted on/inhibited by Chk1/2 Act on phosphate groups |
|
|
|
p19 |
DNA damage pathway
Promotes cell cycle progression Binds and inhibits MDM2's inhibition of p53
Activated by GF's transcription promotion Acts on MDM2 |
|
|
|
GADD45 |
DNA damage pathway
DNA repair pathway
Activated by DNA damage Acts on repair factors |
|
|
|
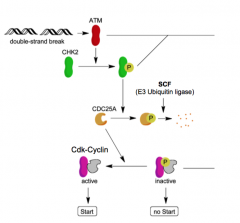
DNA Damage Pathway |
Damage activates ATR/ATM ATM/ATR act on Chk1/2 and p53 Chk1/2 inhibit CDC25 CDC25 acts on phosphate groups p53 acts on p21, MDM2, GADD45, itself p21 acts on CDK/cyclin complexes (CKI) MDM2 inhibits p53 GADD45 activates repair p53 suppresses cell cycle |
|
|
|
EGF Signal Transduction Pathway |
EGF binds EGFR EGFR dimerizes Cytoplasmic tyrosine domains cross-phosphorylated Tyr-P docks Grb2 Grb2 brings SOS via complex SOS activates RAS-GDP, replaces GDP with GTP RAS-GTP docks inactive RAF RAF brought to membrane on CSK platform with inactive KSR KSR activation forms unfolded heterodimer, activates RAF RAF initiates ser/thr kinase phosphorylation cascade Cascade activates MYC, JUN, FOS M, J, F transcribe necessary genes for G1/S initiation |
|
|
|
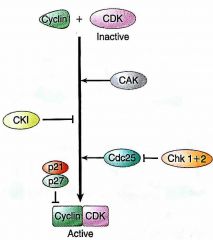
Cyclin/CDK Complex Formation |
CAKs promote CKIs inhibit |
|
|
|
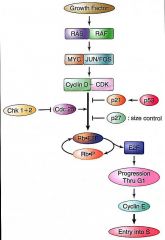
Entry to S-Phase |
Cyclin D/CDK complex phosphorylates Rb Phosphorylated Rb releases bound E2F E2F activates target gene transcription Cyclin E activated in G1, amplifies Rb phosphorylation Right before S-phase, cyclin A synthesized |
|
|
|
Rb |
Tumor suppressor Integrates p53 and cell size signals Normally bound to E2F Phosphorylation required to progress Acted on by Cyclin D/CDK complex |
|
|
|
p27 |
CKI High levels in G0 cells Size control EGFR activation causes p27 reduction |
|
|
|
p16/p19/INK/ARF |
CKIs for Cyclin D |
|
|
|
G1-Phase |
Gap 1 Tightly controlled part of interphase prior to S Length determined by how long it takes to synthesize sufficient cyclin levels to overcome CKI levels |
|
|
|
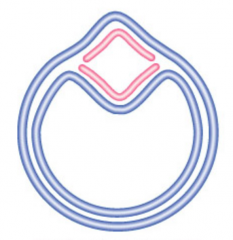
θ Structure
|
Replication bubble in circular DNA |
|
|
|
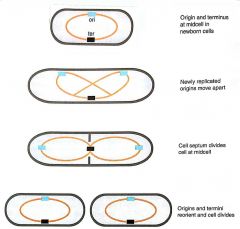
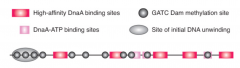
oriC |
Origin of E. coli 245 bp Contains eleven GATC/CTAG repeats that aremethylated on adenine on both strands Must be fully methylated to replicate |
|
|
|
Hemimethylated DNA |
Parental strand of replicated DNA methylated by Dam methylase Cannot initiate replication 13 min delay (from SeqA) before remethylation |
|
|
|
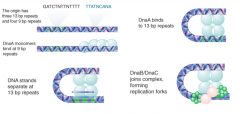
Large Protein Complex |
Sequentially assembled complex on membrane required for initiation at oriC |
|
|
|
DnaA |
Licensing factor necessary for replication Inactivated/destroyed after 1 round of replication Levels low immediately after replication |
|
|
|
DnaA-ATP |
Active form of DnaA Binds to short repeated sequences, forms oligomeric complex that melts DNA |
|
|
|
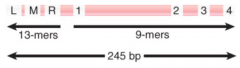
Minimal Origin |
Distance between outside members of 13-mer and 9-mer repeats |
|
|
|
DnaC |
Monomers are loading factors Bind DnaB hexamer DnaB/C complex binds origin of replication |
|
|
|
DnaB |
Hexamer Forms replication fork (with gyrase and SSB) and causes forward movement Initiates each Okazaki frag DnaB/C complex binds origin of replication |
|
|
|
DnaG |
Primase Bound to helicase Interacts w/ DnaB |
|
|
|
Prepriming |
Formation of large protein complex sequentially |
|
|
|
Eukaryotic Replicon |
40-100 kb Individually activated at characteristic times in S-Phase Replicons near one another activated at same time |
|
|
|
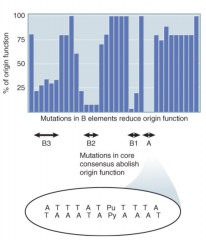
ARS |
Origin of replication in yeast Short A-T sequences w/ essential 11bp sequence (A domain) Binds ORC (6 protein complex) |
|
|
|
Xenopus Egg Experiment |
Nucleus injected into Xenopus egg replicates only once Demonstrates importance of licensing factors, because egg cannot replicate without another factor synthesized Permeabilizing membrane allows more replication, factor enters nucleus post-mitosis |
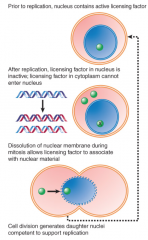
|
|
|
cdc6 |
Unstable licensing factor protein Synthesized only in G1 Binds to ORC Allows MCM to bind |
|
|
|
MCM |
Protein complex Binds to ORC Eukaryotic helicase Mutation in MCM2, 3, 5 prevent replication initiation Some MCM proteins in nucleus throughout cell cycle, others enter only post-mitosis |
|
|
|
cdt1 |
Licensing factor |
|
|
|
Mutations in ubiquination system |
Result in over-replicated DNA Emphasize licensing factor degradation |
|
|
|
Prereplication Complex |
Protein/DNA complex at origin in yeast Required for DNA replication Contains: -ORC -Cdc6 -Cdt1 -MCM |
|
|
|
Complex after Replication Initiation |
cdc6, cdt1, MCM proteins displaced cdc6 degraded, must be resynthesized before next replication round |
|
|
|
Postreplication Complex |
Protein/DNA complex in yeast ORC complex bound to origin |
|
|
|
Replisome |
Multiprotein structure that assembles at replication fork to undertake DNA synthesis Contains DNA polymerase + other enzymes |
|
|
|
Condition Lethal |
Mutation lethal under one set of conditions, but not under a second set Ex: Temperature |
|
|
|
DNA Polymerase I (Pro) |
Gene = polA Major repair enzyme w/ 3' to 5' and 5' to 3' exonucs Single polypep (103kD), can be cleaved into 2 via subtilisin in lab Larger frag = Klenow, contains 3' to 5' exonuc activity Smaller frag = 5' to 3' exonuc activity, excises small groups ~10 bases at a time, provides DNA pol I with ability to start replication at nick in DNA Removes primer and replaces w/ DNA |
|
|
|
DNA Polymerase II (Pro) |
Gene = polB Replication restart |
|
|
|
DNA Polymerase III (Pro) |
Gene = polC Replicase |
|
|
|
DNA Polymerase IV (Pro) |
Gene = dinB Translesion replication |
|
|
|
DNA Polymerase V (Pro) |
Gene = umuD' ₂C Translesion replication |
|
|
|
Proofreading |
Error correction mechanism for DNA synthesis after bases added to chain Fidelity of replication improved x100 |
|
|
|
Processivity |
Ability of enzyme to perform multiple catalytic cycles w/ single template, instead of dissociating after each cycle (Clamp loading/unloading with leading/lagging strands) |
|
|
|
Helicase |
Required to separate DNA strands, needs energy from ATP hydrolysis |
|
|
|
Single-Stranded Binding Proteins |
Maintain the separated strands |
|
|
|
Primers |
Req. to initiate DNA synthesis Needs 3' -OH end Can be: -RNA Primer -Nick in DNA -Priming protein (some viruses) |
|
|
|
DNA Polymerase Holoenzyme |
Consists of subcomplexes -Catalytic core -Dimerization subunit -Processivity component (placed by clamp loader) One catalytic core associated w/ each template strand ε subunit (3' to 5' proofreading exonuc) and θ subunit (stimulates exonuc) associate w/ catalytic core |
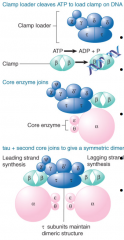
|
|
|
Clamp |
Leading strand core processive b/c clamp keeps it on DNA Lagging strand core dissociates at end of each Okazaki frag and reassembles for next frag Replication fork goes in one direction b/c of lagging strand looping |
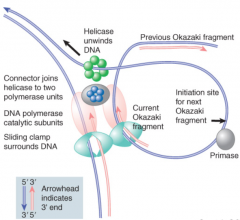
|
|
|
DNA Ligase |
Seals nicks between adjacent nucleotides via enzyme-AMP intermediate Makes bond connecting 3' end of one Okazaki frag to 5' beginning of next frag |
|
|
|
Helicase (Pro vs Euk) |
Pro = DnaB Euk = MCM Complex |
|
|
|
Loading Helicase/Primase (Pro vs Euk) |
Pro = DnaC Euk = cdc6 |
|
|
|
Single-Strand Maintenance (Pro vs Euk) |
Pro = SSB Euk = RPA |
|
|
|
Priming (Pro vs Euk) |
Pro = DnaG Euk = Pol α / Primase |
|
|
|
Sliding Clamp (Pro vs Euk) |
Pro = β Euk = PCNA |
|
|
|
Clamp Loading/ATPase (Pro vs Euk) |
Pro = γδ complex Euk = RFC |
|
|
|
Catalysis (Pro vs Euk) |
Pro = Pol III core Euk = Pol δ (L∆gging) and Pol ε (Lεading) |
|
|
|
Holoenzyme Dimerization (Pro vs Euk) |
Pro = tau Euk = ? |
|
|
|
RNA Removal (Pro vs Euk) |
Pro = Pol I Euk = FEN1 |
|
|
|
Ligation (Pro vs Euk) |
Pro = Ligase Euk = Ligase 1 |
|
|
|
Eukaryote Initiation and Elongation |
One DNA pol α and both complexes of δ or ε Pol α/Primase complex initiates synthesis of both strands ε elongates leading, δ elongates lagging |
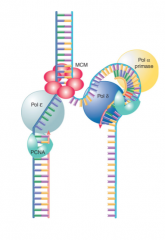
|
|
|
Termination of Replication |
2 E. coli rep forks usually meet ½way around circle ter sites halt rep fork if it goes too far |
|
|
|
Site-Specific Recombination |
Between 2 specific sequences, don't have to be homologous Typically have limited common sequences |
|
|
|
Somatic Recombination |
Occurs in non-germ cells, usually in immune system |
|
|
|
Homologous Recombination Uses |
Generating diversity in meiosis Chromosome Segregation DNA damage and stalled rep fork repair in mitosis |
|
|
|
DNA Polymerase α |
High fidelity replicase Nuclear replication 350 kD tetramer |
|
|
|
DNA Polymerase δ |
High fidelity replicase Lagging strand 250 kD tetramer |
|
|
|
DNA Polymerase ε |
High fidelity replicase Leading strand 350 kD tetramer |
|
|
|
DNA Polymerase γ |
High fidelity replicase Mitochondrial replication 200 kD dimer |
|
|
|
DNA Polymerase β |
High fidelity repair Base excision repair 39 kD monomer |
|
|
|
DNA Polymerase Zeta |
Low fidelity repair Base damage bypass Heteromer |
|
|
|
DNA Polymerase η |
Low fidelity repair Thymine dimer bypass Monomer |
|
|
|
DNA Polymerase ι |
Low fidelity repair Required in meiosis Monomer |
|
|
|
DNA Polymerase κ |
Low fidelity repair Deletion and base substitution Monomer |
|
|
|
Leptotene |
Chromosomes condense, become visible Attach to nuc. envelope Each chromosome replicated, has 2 sister chromatids |
|
|
|
Zygotene |
Chromosomes begin pairing in limited regions Initiation - DNA break induced |
|
|
|
Pachytene |
Synaptonemal complex extends along entire length of paired chromosome Single strands exchange |
|
|
|
Diplotene |
Chromosomes separate, held together by chiasmata Region of exchanged strands is extended |
|
|
|
Diakinesis |
Chromosomes condense, detach from nuc. envelope, chiasmata remain All 4 chromatids become visible DNA cleaved and religated to generate intact products |
|
|
|
Bivalent |
Structure w/ all 4 chromatids (2 per homolog) at start of meiosis |
|
|
|
Synaptonemal Complex |
Protein structure that forms between synapsed homologous chromosomes, important for recombination to occur Brings chromosomes into juxtaposition |
|
|
|
Joint Molecule |
Pair of DNA duplexes connected together through reciprocal exchange of genetic material Forms Holliday junction |
|
|
|
Double-Stranded Break Repair |
1) Make DS break in recipient DNA duplex 2) 5' end resection, exonuc. action generates 3' SS ends that invade donor duplex 3) SS from donor duplex displaces counterpart in recipient, creates D-loop (SS invasion) 4) Exchange generates heteroduplex DNA stretch and 2 Holliday junctions w/ one strand from each parent 5) Recombinant formation depends on if strands involved (in exchange or nonexchange strands) are nicked + ligated during resolution 6) If joints are resolved in opposite ways, genetic crossover produced |
|
|
|
Type I Topoisomerase |
Makes SS breaks Forms covalent bond to one of the broken ends -moves one strand around other, then transfers bound end to other broken end -Bonds conserved -No energy input |
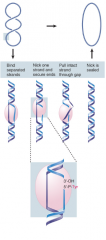
|
|
|
Type II Topoisomerase |
Makes DS breaks Forms covalent bonds to broken ends, then passes duplex DNA region through DS break -ATP required to reseal break |
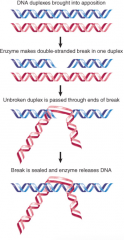
|
|
|
Gyrase |
Enzyme that introduces (-) supercoils into DNA |
|
|
|
Reverse Gyrase |
Enzyme that introduces (+) supercoils into DNA |
|
|
|
Lateral Element |
Structure in synaptonemal complex Forms when pair of sister chromatids condenses onto an axial element |
|
|
|
Central Element |
Structure in synaptonemal complex Located in middle, along which lateral elements of homologous chromosomes align |
|
|
|
Cohesins |
Proteins that form lateral elements |
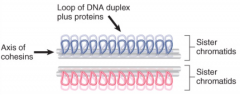
|
|
|
Zip Proteins |
Form transverse central elements |

|
|
|
Lytic State |
λ phage DNA exists as independent, circular molecule in infected bacterium
|
|
|
|
Lysogenic State |
λ phage DNA is integral part of bacterial chromosome
(Part integrated = prophage) |
|
|
|
λ Phage Integration
|
Via recombination between (attachment) attP site on phage and attB site on bacterial chromosome' Needs Int and IHF |
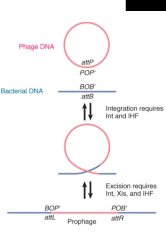
|
|
|
Core Sequence 'O' |
Segment of DNA common to att sites on both λ phage and bacterial genomes Location of recombination event Flanking seq. specific to bacteria or phage (B and B' vs P and P') |
|
|
|
λ Phage Excision
|
Phage excised by recombination between sites at end of linear prophage Needs Xis, Int, IHF |
|
|
|
Int |
λ phage integrase λ phage protein used in integration |
|
|
|
IHF |
Integration Host Factor Bacterial protein used in integration |
|
|
|
Xis |
λ phage protein required in excision |
|
|
|
Integrases |
Proteins that catalyze recombination by similar mech to topoisomerase EXCEPT integrase seals nicked strands from different duplexes Conserves energy using catalytic Tyr in enzyme to break phosphodiester bond and link to broken 3' end No removal/addition of seq. |
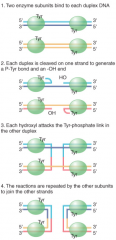
|
|
|
MAT |
Yeast mating type locus Mating type cassette (Allele at MAT = active cassette) Can be MATa or MATα |
|
|
|
HO Allele |
Dominant HO switch mating type frequently Recessive ho mutation at freq of ~10⁻⁶ |
|
|
|
HMLα |
Silent cassette in yeast |
|
|
|
HMRa |
Silent cassette in yeast |
|
|
|
Yeast Mating Type Switch |
Occurs if MATa is replaced by HMRα or MATα is replaced by HMRa Silent cassettes replace active of opposite genotype, have same seqs as corresponding active cassettes (except absence of extreme flanking seqs in HMRa) Initiated by DSB made at MAT locus by HO endonuc |
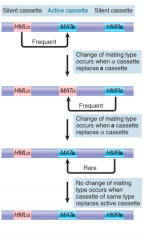
|
|
|
Gene Conversion |
Mating type switching mechanism in yeast Special homologous recombination event Copies info from HMLα or HMRa to active MAT locus |
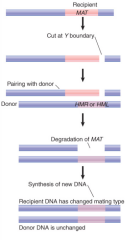
|
|
|
Repair Systems |
Correct DNA errors and chemical modifications of bases -Mismatch pairing -Base conversion |
|
|
|
Thymine Dimers |
Damage to adjacent thymines caused by UV irradiation Blocks replication and transcription Corrected by excision |
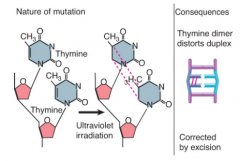
|
|
|
Depurination |
Removal of base from DNA Blocks replication and transcription Corrected by insertion |
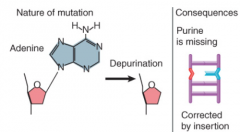
|
|
|
DNA Base Methylation |
Causes distortion of double helix and mispairing at replication Usually pairing to thymine Corrected by dealkylation |
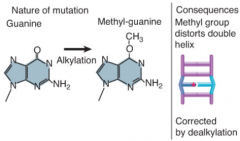
|
|
|
Mismatch Repair |
Corrects recently inserted bases that do not properly pair Preferentially corrects seq of daughter strand New DNA strand synthesized by DNA pol III after excision Pro = based on methylation state Euk = unclear |
|
|
|
Excision Repair |
Correction with direct excision of one DNA strand
Replaced by resynthesis using complementary strand as template |
|
|
|
Photoreactivation |
Direct repair Uses a blue λ light dependent enzyme to split cyclobutane pyrimidine dimers formed by UV Nonmutagenic, acts only on pyrimidine dimers NOT in mammals |
|
|
|
Recombination Repair |
Repair by filling gap in one strand of duplex DNA by using homologous single strand from another duplex AKA postreplication repair Coded for by rec genes in E. coli Preferred mech for DSBs, ensuring no loss of critical genetic info |
|
|
|
Uvr System |
Makes incisions ~12 bases apart flanking damaged DNA Excises DNA between them (DNA pol I, 5'-3') Resynthesis of new DNA (DNA pol I, can be II and III also) UvrAB recognizes damage UvrBC nicks DNA UvrD unwinds marked region |
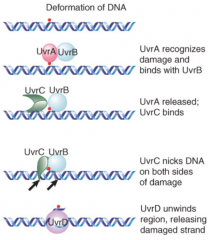
|
|
|
Mfd |
Repair factor Recognizes stalled RNA polymerase Directs Uvr complex to damaged template strand |
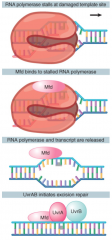
|
|
|
Xeroderma Pigmentosum |
Human disease caused by mutations in any one of several nucleotide excision repair genes Extreme sensitivity to UV, increased skin cancer risk |
|
|
|
Global Genome Repair |
Recognizes damage anywhere in genome Uses XPC as damage recognition mechanism XPE complex detects lesion
XPA binds damaged DNA, recruits helicases XPB and XPD Converges with other pathway DNA excised by 5' XPF and 3' XPG endonucleases and repaired by common proteins |
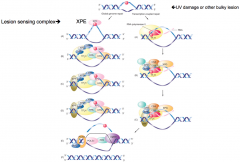
|
|
|
Transcriptional-Coupled Repair |
Repair of transcriptionally active genes Uses RNA pol II as damage recognition mechanism Stalling of RNA pol recruits CSA/B |
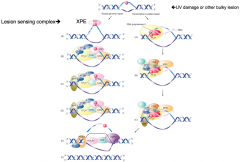
|
|
|
CSA/B |
Cockayne Syndrome proteins used in transcriptional-coupled repair Recruited by RNA polymerase |
|
|
|
XPV |
XP variant Codes for DNA pol η (thymine dimer bypass), allows cell division to proceed with unrepaired damage |
|
|
|
Base Excision Repair |
Direct removal of damaged base Requires glycosylases to recognize uracil and alkylated bases Triggers removal and replacement of polynucleotide stretch Nature of base removal reaction determines which excision repair pathway activated DNA pol δ/ε = long stretch DNA pol β = short stretch |
|
|
|
Glycosylases, Photolyase, Methyl Transferases |
Glycosylases excise bases leaving phosphate backbone intact Photolyases cut out pentose sugar itself Methyl Transferases pop out base, repair, pop back in |
|
|
|
Long-Patch Pathway |
Triggered by base removal by glycosylase APE1 cleaves polynucleotide chain on 5' side Recruits replication complex incl. DNA pol δ/ε -Performs short synthesis rxn extending for 2-10 nucleotides -displaced material removed by endonuc. FEN1 -Ligase 1 seals chain |
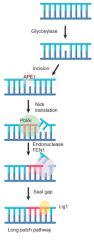
|
|
|
Short-Patch Pathway |
Glycosylase and lyase initially remove base APE1 recruits DNA pol β to replace single nucleotide Nick sealed by ligase XRCC1/ligase 3 |
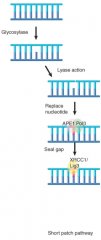
|
|
|
APE1 |
Apurinic/apyrimidinic endonuclease 1 |
|
|
|
Error-Prone Repair and Translesion Synthesis |
Unrepaired DNA causes pol III to stall during replication DNA pol IV or V can synthesize complement to damaged strand (prokaryotes) DNA pol zeta or η can also synthesize complement strand to bypass lesion (eukaryotes) Repair DNA pols often have errors in seq Translesion pols introduce only a short DNA stretch, then replicative pols take over |
|
|
|
Mutator |
Mutation or mutated gene that increases basal level of mutation Often code for proteins involved in repairing damaged DNA |
|
|
|
mut Genes |
Code for mismatched base pair repair system mutH, mutS, mutL, mutY ID'd by mutagenesis screen |
|
|
|
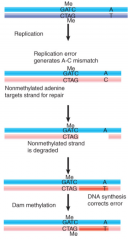
GATC Sequences |
Targets for Dam methylase post-replication Strand lacking methylation at hemimethylated GATC/CTAG usually replaced Mismatch repair removes errors in newly synthesized strand |
|
|
|
dam2 Mutants |
Increased rate of spontaneous mutation Lack of methylation on parental strand makes it difficult to distinguish between daughter and parent, correction of errors decreases |
|
|
|
SeqA |
Binds to hemimethylated DNA to prevent replication immediately after one cycle |
|
|
|
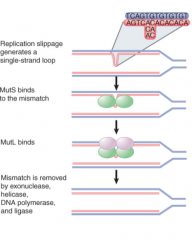
mutS |
Dimer Recognizes mismatch and binds to it Part of mutSL complex |
|
|
|
mutL |
Dimer Binds to mutS |
|
|
|
mutH |
Endonuclease Binds to mutSL after mutSL recognition of GATC seq Cleaves unmethylated strand |
|
|
|
RecJ and Exonuclease VII |
5' to 3' excision of cleaved strand in mismatch repair after cleavage by mutH |
|
|
|
Exonuclease I |
3' to 5' excision of cleaved strand in mismatch repair after cleavage by mutH |
|
|
|
Eukaryotic mutSL Systems |
Repair of mismatches and insertion/deletion loops Do not use DNA methylation to select daughter strand, mechanism not fully understood mutSL homologs interact directly with replication machinery, appear to prefer repair of lagging/nicked strands May utilize gaps between replication bubbles and Okazaki frags to distinguish |
|
|
|
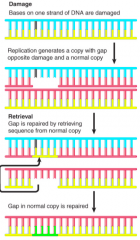
Retrieval System |
E. coli use normal strand of DNA to replace gap left in newly synthesized strand, opposite site of unrepaired damage Use single strand of another duplex (single-strand exchange) Same events must be repeated after every replication cycle unless damage is removed by excision repair system |
|
|
|
Stalled Replication Fork |
Occurs when damaged site or nick in DNA reached Fork can reverse by pairing between the 2 newly synthesized strands DNA pol II required for replication restart, later replaced by DNA pol III |

|
|
|
Rec/Rad |
Proteins for DSB recombination repair Rec = prokaryotes Rad = eukaryotes RecBCD is enzyme in E.coli Has helicase and single stranded nuclease activities RecA binds to ssDNA to form nucleoprotein filament used in strand invasion Rad identified by radiation sensitive phenotypes Rad52 group of genes required for recomb repair in eukaryotes Rad 51 = RecA Rad52, 55, 54 required to form stable Rad51 filament, assist in homology search and strand invasion |
|
|
|
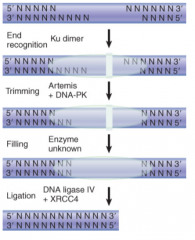
Nonhomologous End Joining (NHEJ) |
Repairs DSBs Used in VDJ recombination for Ig genes Uses Artemis, DNA ligase IV (+XRCC4), and unknown DNA polymerase Not as effective in maintaining sequence integrity as homologous recomb, mutations to these components can render euk cells sensitive to radiation |
|
|
|
Artemis |
Nuclease involved in DSB repair with NHEJ Activated by DNA dependent protein kinase Activated form has endo and exonuc activity |
|
|
|
Ku70/Ku80 |
Heterodimer Senses DNA damage by binding to broken ends Narrow bridge between subunits completely encircles DNA |
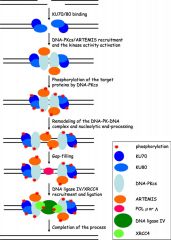
|
|
|
DNA Repair Deficiency |
Causes chromosomal instability and disease Revealed by chromosomal aberrations associated w/ increased mutation rate Leads to increased susceptibility to cancer |
|
|
|
Ataxia Telangiectasia |
DNA repair deficiency disease Failure of cell cycle checkpoint pathway |
|
|
|
Nijmegen Breakage Syndrome |
DNA repair deficiency disease Caused by mutation of repair enzyme |
|
|
|
NSB1 |
Repair factor encoding Nibrin repair enzyme Phosphorylated by ATM in response to irradiation Localizes to site of DNA damage Deficiency of NSB1/ATM leads to -Immunodeficiency -Ionizing radiation sensitivity -Cancer predisposition |
|
|
|
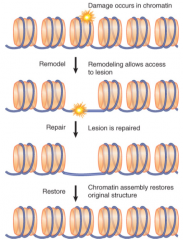
Chromatin Damage |
Must be modified and remodeled before or during repair Original chromatin state must be restored after repair completion H2AX phosphorylation (γ-H2AX) is conserved DSB-dependent modification Recruits chromatin modifying activities and facilitates assembly of repair factors at damage sites Different patterns of histone mods can differentiate stages of repair or pathways |
|
|
|
asf1 and CAF1 |
Remodelers and chaperones required in chromatin repair Reset chromatin structure after repair complete |
|
|
|
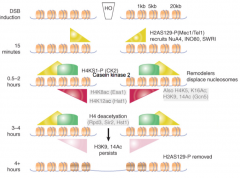
Galactose-Inducible HO DSB |
Histone modification varieties correspond to which kind of repair occurs Mec1/Tel1 phosphorylate H2AX H4 phosphorylated H4 and H3 acetylated H4 deacetylated γ-H2AX dephosphorylated |
|
|
|
Mec1/Tel1 |
Correspond to ATR/ATM respectively Phosphorylate H2AX |
|
|
|
% of Mammalian Genome Composed of Transposons |
~47-50% |
|
|
|
piRNA |
Involved in silencing of transposon activity in germ line cells |
|
|
|
Transposable Element/Transposon |
A DNA sequence able to insert itself or a copy of itself at a new location in the genome, without having any sequence relationship w/ target locus |
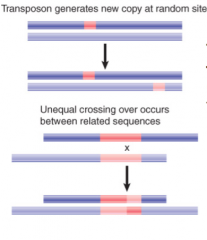
|
|
|
Retrotransposon |
A transposon that mobilizes via RNA intermediate DNA transcribed into RNA, and then reverse transcribed to DNA, which is inserted at new site in genome Does not have infective form Normally refers to retroelements w/ retrovirus-like LTRs and resemble retroviruses |
|
|
|
Insertion Sequence |
Transposon coding for enzymes needed for transposition Flanked by short inverted terminal repeats Target site where transposon is inserted is duplicated during insertion process, forms 2 repeats in direct orientation at ends of transposon (direct repeats) (5 to 9 bp long) |
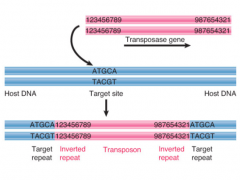
|
|
|
Transposase |
Enzyme activity involved in insertion of transposon at new site |
|
|
|
Composite Transposons/Elements |
Have central region flanked by IS element at each end
Either one or both of IS elements of composite transposon can undertake transposition Can also transpose as a unit |
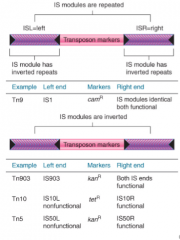
|
|
|
Tn Transposons |
Type of composite transposons Central region has drug resistance marker(s) in addition to transposase Flanked by "arms" consisting of IS elements Arms can be in same or inverted orientation |

|
|
|
Mechanisms of Transposition |
Staggered nicks made in target DNA, transposon joined to protruding ends, gaps filled Starts by forming strand-transfer complex, where transposon is connected to target site through one strand at each end Replicative transposition occurs if complex replicated, nonreplicative if complex repaired Generates crossover, which is converted into cointegrate via replication |
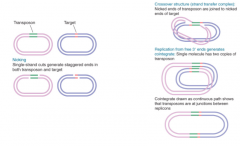
|
|
|
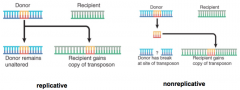
Transposition Mode Determination |
Determined by order of events and nature of connections between transposon and target Replicative -Donor remains unaltered, recipient gains copy of transposon Nonreplicative -Donor has break at site of transposon, recipient gains copy of transposon |
|
|
|
Transposon Resolution |
Catalyzed by enzyme resolvase Uses site-specific recomb between 2 copies of transposon that has been duplicated |
|
|
|
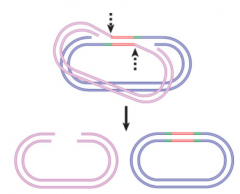
Nonreplicative Transposition |
Results when crossover structure is released by nicking If crossover structure is nicked on unbroken pair of donor strands and target strands on either side of transposon are ligated |
|
|
|
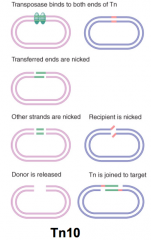
Tn10 |
Transposase functioning as dimer Nonreplicative transposition used First pair of transposon strands are joined to target before second pair are cut |
|
|
|
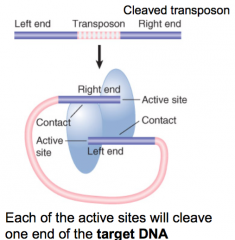
Tn5 |
Transposase functioning as dimer Nonreplicative transposition used All four strands cut before joining to target |
|
|
|
Transposon Excision |
Homologous recomb between multiple transposon copies causes rearrangement of host DNA Homologous recomb between repeats of transposon may lead to precise or imprecise excision Reciprocal recomb between direct repeats excises material between them Reciprocal recomb between inverted repeats inverts region between them Each product of recomb has one copy of direct repeat |
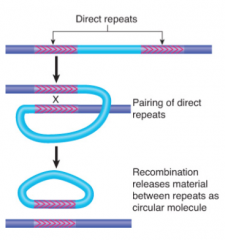
|
|
|
Hybrid Dysgenesis |
Caused by transposition of P elements Inability of certain strains of fruit fly to interbreed b/c hybrids sterile, but may be phenotypically normal Asymmetrical, induced by: P-male x M-female NOT M-Male x P-Female Activation of transposition causes P element insertion at sites rendering sterility Determined by interactions between P elements in genome and 66kD repressor in cytotype |
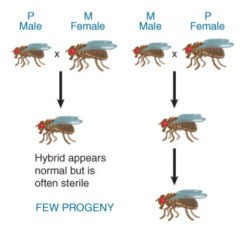
|
|
|
P Elements
|
Transposons carried in P strains of fruit fly, not in M strains Has 4 exons First 3 spliced together in somatic expression All 4 spliced together in germline expression P elements actiated in germline of P-Male x M-Female b/c tissue-specific splicing removes 1 intron, which generates coding seq for transposase |
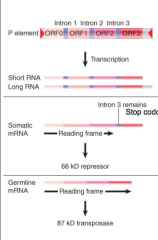
|
|
|
P Element Repressor |
Inherited maternally in cytoplasm Presence explains why M-Male x P-Female remains fertile May consist of piRNAs derived from P element transcripts that target destruction of other P element transcripts |
|
|
|
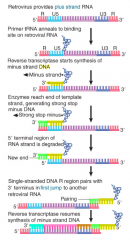
Retrovirus |
Virus that uses RNA as genetic material Has 2 copies of genome (2 SS RNAs) Generates provirus via reverse transcription of genome, additional retroviral genomes can be generated via provirus transcription Has 3 genes: -gag -pol -env |
|
|
|
Provirus |
Integrated DS DNA sequence from virus |
|
|
|
Reverse Transcriptase |
Enzyme using SS RNA as template to synthesize complementary DNA strand |
|
|
|
Integrase
|
Enzyme responsible for site specific recomb that inserts one molecule of DNA into another |
|
|
|
gag |
Viral gene ~2000bp Translated from full length transcript of genome Codes for MA, CA, NC (matrix, capsid, nucleocapsid) |
|
|
|
pol |
Viral gene ~2900 bp Translated from full length transcript of genome Translation requires readthrough or frameshift by ribosome Codes for PR, RT, IN (protease, reverse transcriptase, integrase) |
|
|
|
env |
Viral gene ~1800 bp Translated from separate mRNA generated by splicing Codes for SU and TM (surface protein, transmembrane) |
|
|
|
Polyproteins |
Genes in retrovirus expressed as polyproteins which are processed by proteases to give multiple individual protein products |
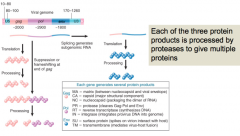
|
|
|
HIV Transport Mechanism |
Budding from plasma membrane of infected cell |
|
|
|
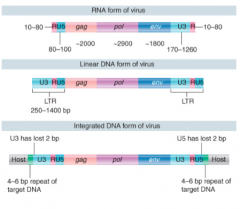
R Sequence (Retroviral) |
Short repeated sequence at each end of viral RNA, giving R-U5 and U3-R sequences |
|
|
|
Plus-Strand Virus |
Virus with a SS nucleic acid genome whose sequence directly codes for protein products |
|
|
|
Minus-Strand DNA |
SS DNA sequence complementary to the viral RNA genome of a plus-strand virus |
|
|
|
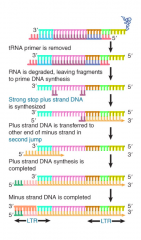
Plus-Strand DNA |
Strand of the duplex sequence representing a retrovirus that has the same sequence as that of the RNA |
|
|
|
Retroviral RNA Conversion |
RT starts synthesis when tRNA primer binds to site 100-200 bases from 5' end When enzyme reaches end, 5' terminal bases of RNA degraded, exposing 3' end of DNA product Exposed 3' end of DNA product base pairs w/ 3' terminus of another RNA genome Synthesis continues, generating project w/ repetition of 5' and 3' regions, giving each end structure of U3-R-U5 |
|
|
|
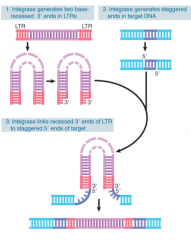
Retroviral RNA Integration |
Integrase only viral protein needed After synthesis, each LTR loses 2 bp and is inserted between 4-6 bp repeats of target DNA Organization of proviral DNA in a chromosome is same as a transposon, w/ provirus flanked by short direct repeats of a sequence at a target site Linear DNA inserted directly into host chromosome by integrase |
|