Reading...
![]()
Play button
![]()
Play button
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
189 Cards in this Set
- Front
- Back
|
Alanine
|

|
|
|
Asparagine
|

|
|
|
Arginine
|

|
|
|
L-Glutamine
|

|
|
|
L-Methionine
|

|
|
|
L-Tyrosine
|

|
|
|
L-histidine
|

|
|
|
L-lysine
|

|
|
|
L-phenylalanine
|

|
|
|
L-proline
|

|
|
|
L-serine
|

|
|
|
L-threonine
|

|
|
|
L-valine
|

|
|
|
Methionine
|

|
|
|
Tyrosine
|

|
|
|
General Structure of Amino Acid
|

|
|
|
aspartic acid
|

|
|
|
cysteine
|

|
|
|
glycine
|

|
|
|
isoleucine
|

|
|
|
leucine
|

|
|
|
phenylalanine
|

|
|
|
proline
|

|
|
|
serine
|

|
|
|
threonine
|

|
|
|
tryptophan
|

|
|
|
4-hydroxyproline
|

found in collagen, formation requires Vitamin C
Scurvy is a disease that results from inadequate Vitamin C; it's a disease of connective tissue |
|
|
Functions of amino acids(4)
|
A) Metabolic energy (4 kcal/g), production or storage in fat
B) Replenish intermediates in central pathways during fasting C) Precursors to neurotransmitters and hormones D) Basic building blocks for proteins |
|
|
Stereochemistry of Amino acids: What handedness are amino acids in proteins?
|
L
|
|
|
Nonpolar, aliphatic amino acids
|
Glycine
Alanine Proline Valine Leucine Isoleucine Methionine |
|
|
Wavelength that aromatic amino acids absorb light and is often used to detect and measure protein concentrations in solution
|
280 nm
|
|
|
Aromatic amino acids
|
Phenylalanine
Tyrosine Tryptophan |
|
|
Polar, uncharged amino acids
|
Serine
Threonine Asparagine Glutamine Tryptophan Tyrosine |
|
|
Sulfur-containing amino acids
|
Cysteine (Cys, C)
Methionine (Met, M) |
|
|
Aliphatic alcohols
|
Serine (Ser, S) - R group is CH2-OH
Threonine (Thr, T) - R group is CHOH-CH3 |
|
|
Basic side chains (positive charge at neutral pH)
|
Histidine (His, H) - R group is imidazole
Lysine (Lys, K) - R-group is CH2-CH2-CH2-CH2-NH3+ Arginine (Arg, R) - R-group is CH2-CH2-CH2-NH-C(=NH2)-NH2 |
|
|
Positively charged amino acids
|
Lysine
Arginine Histidine (basic side chains) |
|
|
Carboxyl side chains
Negatively charged at physiological pH |
Aspartate (Asp, D) R group is CH2-CO2-
Glutamate (Glu, E) - R-group is CH2-CH2-CO2- |
|
|
Amide side chains (of acidic amino acids)
|
Asparagine (Asn, N) - R group is CH2-CO-NH2
Glutamine (Gln, Q) R-group is CH2-CH2-CO-NH2 |
|
|
Nonpolar amino acids
|
Ala, Cys, Gly, Met, Leu, Phe, Pro, Trp, Val, Ile
|
|
|
Polar, uncharged amino acids
|
Asn, Gln, Ser, Thr, Trp, Tyr
|
|
|
Polar, charged amino acids
|
Glu, Arg, Lys, His, Asp
|
|
|
Gamma carboxyglutamate
|

Formation requires Vitamin K
Occurs in blood-clotting protein, prothrombin |
|
|
Coumadin
|
vitamin K antagonist
impairs formation of gamma carboxyglutamate |
|
|
Selenocysteine
|

Plays role in proteins involved in antioxidative mechanisms. Selenium is an essential micronutrient but toxic in higher concentrations
|
|
|
Essential Amino Acids
|
PVT TIM HALL
Phe, Val, Thr, Trp, Iso, Met, His, Arg, Leu, Lys |
|
|
Peptide bond formation
|

|
|
|
Properties of peptide bond influencing structure of proteins
|
1) Because of resonance forms there is significant double bond character, preventing rotation about C-N bond, resulting in rigid, planar structure
2) With a few impt exceptions, peptide bonds are formed in trans configuration, with alpha carbons on opposite sides of the peptide bond |
|
|
Cystine: what happens when cysteine residues interact?
|
Under oxidizing conditions, they form a covalent disulfide linkage, referred to as Cystine.
Disulfide bonds usually form between cysteines that are distant in the linear sequence and contribute to folding behavior of the protein and stability. |
|
|
Disulfide bond formation/mechanism
|

|
|
|
Which amino acid has a alpha imino group instead of amino?
|
Proline
|
|
|
Ion product of water
|
Kw = [H+][OH-] = 1.0 X 10^-14 M^2
|
|
|
ph : Definition
|
-log [H+]
|
|
|
pH of blood
|
7.4
|
|
|
Equilibrium constant : Ka defn
|
Ka = [H+][A-] / [HA]
|
|
|
Do strong acids have larger or small dissociation constants?
|
Larger
|
|
|
Henderson-Hasselbach equation
|
pH = pKa + log ([A-] / [HA])
Derived by taking -log of Ka and solving for -log[H+] |
|
|
How many pKa's are there in amino acid?
|
Depends. At least two (for carboxy and amino end), and however many more acidic or basic groups on the R group.
|
|
|
Primary Structure of Proteins
|
Amino Acid sequence
Protein rich in hydrophobic a.a.s MAY be a membrane protein, but difficult to infer too much from primary structure |
|
|
Prosthetic group
|
A cofactor emphasizes the nature of its binding to a protein (tight or covalent) and thus refers to a structural property
E.g., carbohydrates, phosphate groups, heme, Iron |
|
|
Secondary Structure of Proteins
|
Energetically favorable, Repeating pattern of structure defined by spatial relationships between amino acid residues close to each other in amino acid sequence.
Generally stabilized by non-covalent interactions (h bonding, hydrophobic) Ex: alpha helix, beta sheet |
|
|
Alpha helix
|
Spiral-shaped conformation of polypeptide backbone with side chains directed OUTWARD
Stabilized by hydrogen bonding between alpha amino hydrogen of peptide bond, and carbonyl oxygen four residues distant in direction of the amino terminus. |
|
|
What stabilizes the alpha helix?
|
1) H-bonding between peptide bond alpha amino hydrogen and carbonyl oxygen four residues away (in direction of amino terminus)
2) Electrostatic interactions between oppositely charged R groups 3-4 residues away 3) Negatively charged residues at amino end (because of dipole with partial positive charge at amino end); likewise positively charged amino acids at carboxyl end |
|
|
What things can destabilize the alpha helix?
|
1) Identical charges on adjacent residues
2) Bulky R groups in close proximity 3) Negatively charged amino acids at carboxy end (because of dipole with partial negative charge at carboxyl end), likewise Positively charged amino acids at amino end. |
|
|
Which amino acids do NOT work well in alpha helix?
|
1) Proline - rigid ring does not permit rotation around the alpha carbon - nitrogen bond, and doesn't h-bond.
2) Glycine - contributes greater flexibility, favors Beta sheets |
|
|
β-sheets
|
Composed of 2 or more different regions of stretches of at least 5-10 amino acids.
Stabilized by H-bonding between amide nitrogens and carbonyl carbons either parallel or antiparallel. In parallel sheets adjacent peptide chains proceed in the same direction (i.e. the direction of N-terminal to C-terminal ends is the same), whereas, in antiparallel sheets adjacent chains are aligned in opposite directions. |
|
|
β-turn
|
Where polypeptide chain changes direction! Permits folding.
Composed of 4 aa residues, contains Gly (unusual flexbility) and Pro (forms peptide bond with cis conformation) |
|
|
Tertiary Protein Structure
|
complete three-dimensional structure of the polypeptide units of a given protein
|
|
|
Quaternary Protein Structure
|
Structural arrangement resulting from interaction with other polypeptides (not all proteins have)
|
|
|
Characteristics of properly folded protein
|
Hydrophobic residues in core
Ionic and polar residues on surface |
|
|
How many residues in one turn of alpha helix?
|
3.4
|
|
|
Isoelectric point (pI)
|
Point at which molecule has a net neutral charge. May be in zwitterion form.
Generally it's the average of both pKas When pH > pI, net charge is negative. When pH < pI, net charge is positive. |
|
|
What is pKa?
|
Point at which the weak acid is half ionized.
When pH = pKa, half is ionized |
|
|
Buffering region
|
+/- 1 pH unit of pKa
Area where adding more base will not make huge difference in pH |
|
|
What is the general pKa of the FIRST acidic group (alpha carboxy) of amino acid?
|
Around 2
|
|
|
What is the general pKa of the alpha amino group of an amino acid?
|
Around 9-10
|
|
|
When is an amino acid LEAST soluble?
|
At pI - it has no net charge so it's hard to get it out (can't bind to ion exchange column etc)
|
|
|
Site of ribosomal RNA synthesis
|
Nucleolus
|
|
|
Where are chromosomes located?
|
Nucleus (duh)
|
|
|
Where are ribosomes located?
|
in Rough ER and in cytosol
|
|
|
Nucleotides - what do they contain?
|
Base + sugar + phosphate group
|
|
|
Nucleosides - what do they contain?
|
Base + sugar
|
|
|
Thymine
|

|
|
|
Adenine
|

|
|
|
Guanine
|

|
|
|
Thymine
|

|
|
|
Uracil
|

|
|
|
Cytosine
|

|
|
|
What does cytosine deaminate to?
|
Uracil. Uracil IS thymine without the methyl group.
|
|
|
Nucleoside nomenclature.
|
NucleoSIDEs end with -SINE or -DINE
Adenosine, Guanosine Cytidine, Thymidine, Uridine |
|
|
Nucleotide nomenclature
|
NucleoTIDES end with -YLATE (no exceptions)
Adenylate, Guanylate, Cytidylate, thymidylate, uridylate |
|
|
ATP structure
|

|
|
|
Which phosphate is ATP is the best leaving group?
|
Gamma (the one on the end). Liberates lots of deltaG
|
|
|
Cyclic AMP
|

Note Phosphate is connected at 3' and 5' position.
|
|
|
Cyclic GMP
|

Note Phosphate is connected at 3' and 5' position.
|
|
|
What enzyme converts ATP to cAMP?
|
adenylyl cyclase
|
|
|
Name of the type of bonds that link nucleotides?
|
Phosphodiester
|
|
|
Molecules required for de novo synthesis of nucleotide
|
1) NH3 (released from glutamine)
2) CO2 (directly in form of bicarbonate) 3) Certain amino acids 4) Ribose 5-phosphate |
|
|
Structure of 5-phosphoribosyl-1-pyrophosphate (PRPP)
|

|
|
|
What's the reaction for how PRPP gets made?
|
Ribose 5-Pi + ATP -> 5-PRPP + AMP
|
|
|
Where do atoms in the purine come from?
|
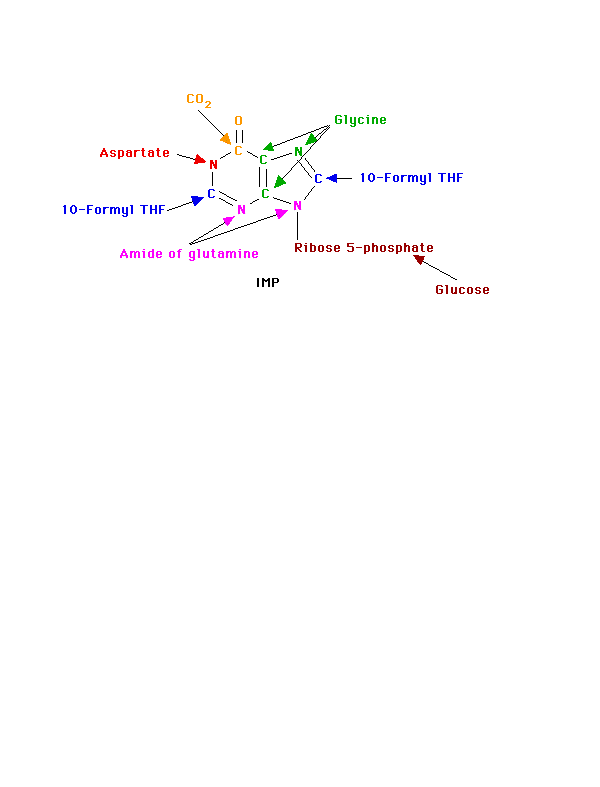
|
|
|
What are the 2 types of pathways leading to nucleoTide biosynthesis?
|
1) De novo synthesis
2) Salvage pathways (from nucleosides and free bases) |
|
|
Where does the sugar component of purines ULTIMATELY come from? (come from first)
|
Glucose
|
|
|
Name 3 roles for PRPP
|
1)purine and pyrimidine nucleotide synthesis
2)salvage pathways 3)NAD and NADP formation. The enzyme is heavily controlled by a variety of compounds (di- and tri-phosphates, 2,3-DPG), presumably to try to match the synthesis of PRPP to a need for the products in which it ultimately appears. |
|
|
1st COMMITTED step in PURINE synthesis (also rate-limiting step)
|
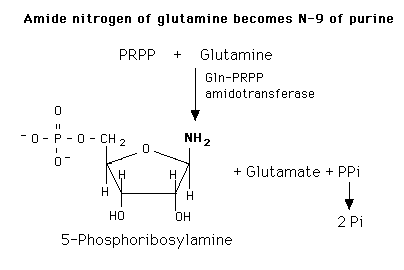
When C1 of PRPP gets attacked by NH2 from glutamine
|
|
|
2nd step in purine synthesis
|
Addition of 2 carbons and 1 nitrogen from glycine (5,4,7)
|
|
|
Hypoxanthine is a base. What is it called as a nucleoside and -tide?
|
Side = inoSine
Tide = inosinATE |
|
|
Describe feedback inhibition for committed step in purine synthesis (creation of 5-Phosphoribosylamine from PRPP)
|
*AMP, GMP, or IMP alone will inhibit the amidotransferase
*AMP + GMP or AMP + IMP together act synergistically. *Also controlled by intercellular concentration of glutamine and PRPP This is a fine control and probably the major factor in minute by minute regulation of the enzyme. The nucleotides inhibit the enzyme by causing the small active molecules to aggregate to larger inactive molecules |
|
|
Conversion of IMP (inosinate) to AMP (adenylate)
|
Requires GTP (->GDP) and aspartate (enzyme is adenylsuccinate synthetase).
A fumarate comes off, and there is AMP! Feedback inhibition: the presence of AMP inhibits conversion of IMP -> AMP |
|
|
Conversion of IMP (inosinate) to GMP (guanylate)
|
First C2 gets oxidized to C=O (conversion of NAD+ to NADH.) Then it's xanthine.
Then ATP is hydrolyzed similarly to conversion of IMP to AMP, except PPi is released Feedback inhibition: the presence of GMP inhibits conversion of IMP -> GMP |
|
|
If both AMP and GMP are present in adequate amounts, where is the de novo pathway turned off?
|
amidotransferase step
(the committed step where glutamine NH2 gets added to PRPP) |
|
|
How are nucleoside diphosphates (NDPs) and triphosphate (NTPs)made from nucleoside monophosphates (NMPs?)
|
1)Base-specific NMP kinases (eg. adenylate kinase, guanylate kinase) add 2nd phosphate
(ATP is source of transferred phosphate) 2) nonspecific NDP kinase adds the second phosphate to make NTPs. |
|
|
Regulatory mechanism for Purine nucleotide synthesis
|
1) Most important is feedback inhibition of committed step, PRPP to 5phosphoribosylamine. AMP, GMP, and IMP presence inhibit.
2) Short feedback loops: AMP presence inhibits IMP formation to AMP, and GMP presence inhibits IMP formation to XMP (XMP would become GMP) 3) Balancing ATP and GTP production |
|
|
Salvage pathways of nucleotide biosynthesis: what gets recycled?
|
Nucleosides
Free bases |
|
|
Sources of atoms in pyrimidine ring
|
Glutamine
CO2 Aspartate |
|
|
What is the COMMITTED step for PYRIMIDINE synthesis?
|
Carbamoyl phosphate + aspartate --> N-CARBAMOYLASPARTATE
|
|
|
Structure of carbamoyl phosphate
|

|
|
|
Where does de novo pyrimidine biosynthesis happen in the cell?
|
cytosol
|
|
|
How is pyrimidine de novo biosynthesis regulated?
|
Feedback inhibition - presence of CTP inhibits committed step.
|
|
|
What kind of proteins are fibrous?
|
Generally, structural (connective tissue)
|
|
|
Fibrous protein characteristics
|
Generally consist of one type of secondary structure repeated, adapted for stuctural function
All insoluble in water |
|
|
Keratin: structure
|
Alpha helix
then coiled onto another alpha helix then attached in protofilaments Then attached in protofibrils cross-linked by disulfide bonds HAIR |
|
|
Collagen: structure and purpose
|
Purpose: connective tissue
Structure: Repeating tri-peptide unit Gly-X-Y; Left-handed helix, Then it associates with three strands to form Trimer (which is right handed) The 3stranded helix is held in superhelix via TIGHT JUNCTIONS. Glycine is required for this b/c of small size |
|
|
What kind of cross-links reinforce collagen fibrils?
|
Cross-links between Lys and HyLys (requires Vitamin C to form HyLys)
|
|
|
Scurvy: Causes and Symptoms
|
Cause: Vitamin C deficiency, prevents formation of HyLys and HyPro. General degradation of connective tissue.
Symptoms: multiple, but include small hemorrhages causes by fragile blood vessels, tooth loss, eventually heart failure |
|
|
Globular proteins : do they have one secondary structure?
|
No, they generally have multiple secondary structures like alpha helices and beta sheets and beta turns
|
|
|
The heme group in myoglobin binds oxygen. Where on hemoglobin is it located and why?
|
It's located near the surface but shielded somewhat (a crevice). It must be maintained in reduced Fe2+ state in order to reversibly bind Oxygen, and the outside is an oxidizing environment.
|
|
|
Structurally similar proteins have similar amino acids sequences: True or False?
|
FALSE! They can have similar motifs made of different amino acids combos. Three-dimensional structure is more important (generally) than primary amino acid sequence.
|
|
|
Protein Domains
|
Distinct globular regions in globular proteins, tend to retain characteristic 3d structures even when separated from rest of protein. May be associated with a particular function or property of the protein (ex. substrate binding, generation of immune response)
|
|
|
Conditionally essential amino acids
|
Histidine, Arginine
(only a few examples) Can't be produced under extreme stress |
|
|
Prions
|
Improperly folded proteins that influence other proteins to misfold. They're infectious, very difficult to destroy, resistant to heat and proteases. Detergent "kills" them.
|
|
|
How to find pI when there are 3 pKas?
|
Average either acidic or basic pKas depending on whether R group is acidic or basic
|
|
|
3 ways to separate protein based on solubility
|
1) Adjust pH
2) Adjusting salt ([NH4]2SO4) concentration. Different proteins precipitate at different ionic strengths. Presence of other ions weakens electrostatic and H bonding 3) Increase temp. Different proteins vary in stability to heat. |
|
|
Column Chromatography
|
Mixture to be separated is poured through a series of porous particles (stationary phase) in a solvent, usually water (mobile phase).
Proteins partition between stationary and mobile phase based on some property (e.g., size, charge, presence of some functional group) |
|
|
Ion Exchange Chromatography
|
Stationary phase is anionic resin with positively charged group or cationic resin with negatively charged group. Mobile phase is buffer.
|
|
|
Why might one opt for eluting an ion exchange column with an ionic strength gradient rather than a pH gradient?
|
pH changes can damage the protein
|
|
|
Size exclusion chromatography
|
Type of column chromatography separates based on size. Small proteins get stuck in porous beads, while large proteins can't fit in the beads and slide right out.
|
|
|
Affinity chromatography
|
Type of column chromatography , similar to size exclusion, but with addition of covalently attached ligand for which the protein of interest has a specific binding affinity. Other proteins then pass through the column. The protein of interest is eluted by adding unbound ligand to the mobile phase.
|
|
|
Electrophoresis
|
protein characterization technique based on mobility of charged particle in electric field.
|
|
|
SDS-PAGE
|
Polyacrylamide gel electrophoresis with Sodium dodecyl sulfate (a detergent) used to denature protein, bind them in constant ratio to molecular weights and causes them to assume similar shapes. Electrophoretic mobilities are roughly proportional to monomeric molecular weights
|
|
|
Western blotting
|
SDS-PAGE with additional step of blotting proteins on membrane and reacting them with enzyme linked antibody recognizing only protein of interest.
|
|
|
Isoelectric focusing
|
Ampholytes (mix of organic acids and bases) added to electrophoresis gel and subjected to an electric field until a pH gradient is formed across the gel. Protein migrates to position where pH generated by ampholyte gradient matched pI of protein.
|
|
|
Two-dimensional gel electrophoresis
|
1) Proteins first separated by electrophoresis on isoelectric focusing gel.
2) This gel then placed sideways on an SDS PAGE and proteins are subjected. Results in proteins separated by both pI values and molecular weights. |
|
|
Direct / overlapping protein (amino acid) sequencing
|
Initial cleavage of peptide bonds with an amino acid specific chemical reagent or protelytic enzyme followed by sequencing of resulting small peptides. Use of different reagents on multiple aliquots of the protein generates a panel of peptides with overlapping sequences, and the amino acid composition can be deduced.
|
|
|
Indirect / translation of nucleotide sequences for protein sequencing
|
It is easier to sequence nucleotides in DNA than in protein. Thus, amino acid sequences of many proteins are deduced from corresponding nucleotide sequences that encode them.
|
|
|
Mass spectrometry: how is it used to sequence proteins?
|
Electrospray Ionization involves introduction of intact charged protein molecules in gas phase into mass spectrometer which analyzes their mass to charge ratio and permits highly accurate determination of their masses.
The masses are detected so precisely that the primary sequence of a protein can be deduced. BONUS: yields info about post-translational processing |
|
|
X-ray crystallography: how is it used to characterize 3d shape of protein?
|
way of deducing 3d shape of protein.
Derives coordinates of individual atoms in a protein crystal from Xray scattering patterns and reconstructs the 3d structure from the coordinates. requires uniform crystal and generates static model (snapshot). Info it provides is only as good as the assumption the crystal structure is similar to functional protein. |
|
|
Nuclear magnetic resonance: how is it used to characterize 3d shape of protein?
|
Analyzes chemical shifts to id and determine proximities of various nuclei in a protein. Computations are very complex and requires lots of protein. But it does produce dynamic data from proteins in solution.
|
|
|
Homology modeling: how is it used to characterize 3d shape of protein?
|
Use of a protein database of xray crystal structures to compare with new protein sequence. The more similar are proteins in sequence, the more likely they'll be similar in higher orders. Computer programs can construct a tentative 3d model based on known structures of peptides with similar sequences.
|
|
|
Mapping: how is it used to characterize 3d shape of protein?
|
Chemical reagents (cross-linkers) or spectral techniques (fluorescence resonance energy transfer) are used to determine proximity of specific residues or group within a protein.
|
|
|
What's special about thymine de novo synthesis?
|
It can't be made from the ribonucleotide form...must be made from deoxyribonucleotide
|
|
|
Thymidylate synthetase
|
Adds methyl group to 5' carbon of dUMP to create dTMP. Requires folate derivative as contributor of methyl group.
|
|
|
5-fluorouracil as chemotherapy agent: how does it work?
|
Inhibits thymidine synthetase from making thymine from uracil.
|
|
|
Methotrexate as chemotherapy agent: how does it work?
|
Inhibits dihydrofolate reductase: looks like dihydrofolate but inhibits production of dTMP.
|
|
|
thymidine kinase
|
The salvage pathway to dTTP synthesis involves the enzyme thymidine kinase which can use either thymidine or deoxyuridine as substrate:
thymidine + ATP <——> TMP + ADP deoxyuridine + ATP <——> dUMP + ADP The activity of thymidine kinase (one of the various deoxyribonucleotide kinases) is unique in that it fluctuates with the cell cycle, rising to peak activity during the phase of DNA synthesis; it is inhibited by dTTP. |
|
|
PRPP synthetase
|
makes PRPP from Ribose-5-phosphate and ATP, releases AMP.
|
|
|
RIBONUCLEOTIDE REDUCTASE
|
CONVERTS NDP -> dNDP. NOTE THAT HIGH [ ]s of dATP is a STRONG INHIBITOR
|
|
|
Glutamine PRPP amidotransferase
|
Adds N to PRPP in rate limiting, committed step. The N comes from glutamine.
It's inhibited by presence of AMP and GMP |
|
|
dihydrofolate reductase (DHFR)
|
Involved in conversion of dUMP to dTMP. (along with thymidylate synthase)
Tetrahydrofolate (THF) is regenerated from the dihydrofolate (DHF) product of the thymidylate synthase reaction by the action of dihydrofolate reductase (DHFR), an enzyme that requires NADPH. |
|
|
Degradation of pyrimidines: Cytosine
|
Cytosine deaminated to uracil which is hydrolyzed to CO2, NH4+ (enters urea cycle), and beta alanine
Occurs primarily in liver |
|
|
Degradation of pyrimidines: Thymine
|
Thymine is hydrolyzed to CO2, NH4+ (enters urea cycle), and beta aminoisobutryate
|
|
|
Is the failure to properly degrade pyrimidines causes of any known health problems?
|
no
|
|
|
ribonucleotide reductase
|
Converts NDP -> dNDP. dATP Strongly inhibits
|
|
|
adenosine deaminase
|
converts adenosine -> inosine
which is ultimately degraded to uric acid Deficiency in this would lead to reduced degradation and increased levels of dATP, which lead to decreased DNA synthesis. Causes Severe Combined Immune Deficiency (SCID) syndrome |
|
|
xanthine oxidase
|
Forms xanthine and uric acid.
Allopurinol (gout treatment) inhibits, so excreted products are more water soluble hypoxanthine and xanthine |
|
|
Degradation of purines all converge to what?
|
xanthine, then to uric acid
|
|
|
Purine bases: salvage pathway
|
Involve re-attchment of base to PRPP by
1)adenosine phosphoribosyltransferase (APRT) 2)hypoxanthine-guanosine phosphoribosyltransferase (HGPRT) |
|
|
adenosine phosphoribosyltransferase (APRT)
|
Involved in purine salvage pathway. Adds Adenine to PRPP resulting in AMP and PPi.
|
|
|
hypoxanthine-guanosine phosphoribosyltransferase (HGPRT)
|
Involved in purine salvage pathway. Adds Hypoxanthine/Guanine to PRPP resulting in IMP/GMP + PPi.
Deficiency in enzyme -->Lesch-Nyhan |
|
|
Is it easier for mammalian cells to salvage free pyrimidine bases or free purine bases?
|
Purine
|
|
|
Lesch-Nyhan syndrome
|
Disease associated with deficits of purine degradation/salvage pathway.
Caused by less salvage and more degradation because deficits hypoxanthine-guanosine phosphoribosyltransferase (HGPRT) |
|
|
severe combined immunodeficiency disease (SCID)
|
caused by a deficiency in the enzyme adenosine deaminase (ADA). This is the enzyme responsible for converting adenosine to inosine in the catabolism of the purines. This deficiency selectively leads to a destruction of B and T lymphocytes, the cells that mount immune responses. In the absence of ADA, deoxyadenosine is phosphorylated to yield levels of dATP that are 50-fold higher than normal. The levels are especially high in lymphocytes, which have abundant amounts of the salvage enzymes, including nucleoside kinases. High concentrations of dATP inhibit ribonucleotide reductase (see below), thereby preventing other dNTPs from being produced. The net effect is to inhibit DNA synthesis.
|
|
|
What do increased concentrations of dATP do?
|
Powerfully inhibit ribonucleotide reductase, causing decreased production of all other dNTPs, severely inhibiting replication of T lymphocytes
|
|
|
Percentage of human DNA representing genes, and percentage representing exons
|
about 30%, and about 1.5%
|
|
|
strand labels: template strand
|
the "anti-sense" strand that is used as a template to transcribe rna. So the rna will be COMPLEMENTARY to it.
Talked about 3' to 5' |
|
|
strand labels: nontemplate strand
|
the "sense" strand that the RNA, transcribed from the template strand, will be identical to (except Ts substituted for Us)
Talked about 5' to 3' |
|
|
What are the 3 basic things needed to transcribe RNA?
|
1) DNA template
2) NTPs (nucleoside triphosphates) 3) RNA polymerase |
|
|
What is the leaving group of newly synthesized RNA molecules?
|
Two phosphate groups from the NTPs...they're incorporated with only one phosphate group
|
|
|
Which direction is "reading" of template strand?
|
3' - 5'
So the gene (sense strand) is transcribed 5' - 3' |
|
|
Major classes of RNA
|
ribosomal rRNA
transfer tRNA messenger mRNA other RNA (like small nuclear RNAs) |
|
|
Prokaryotic RNA polymerase: how many?
|
one, synthesizes all classes of RNAs
|
|
|
RNA polymerase holoenzyme
|
Core RNA polymerase (5 subunits) plus sigma subunit. Sigma is required to bind to promoter. Sigma falls off when transcription starts
|
|
|
Eurkaryotic RNA polymerases: What do each of them do?
|
RNA pol 1: rRNA
RNA pol 2: mRNA and most SNURPS RNA pol 3: tRNA and 5S rRNA Mitochondrial RNA pol: makes all mitochondrial RNAs. |
|
|
Promoters: defn
|
specific DNA sequences around which RNA synthesis begins
There's usually one at -10 (TATA box) and -35, referring to the non-template strand |
|
|
UP element
|
A promoter element characteristic of certain strong promoter regions. Consists of an AT rich region. Tends to be found in frequently transcribed genes.
|
|
|
Sigma subunit of RNA polymerase
|
The binding of sigma subunit is (almost) required to initiate transcription. When it's bound to the core RNA pol, the holoenzyme has higher affinity for promoter regions. The tighter the sigma subunit binds, the more commonly the gene gets transcribed.
|
|
|
If the unique feature of DNA is on the INSIDE (outside is same Phosphate groups), how do RNA polymerases (and gene regulatory proteins) recognize discrete sequences?
|
Different base pairs can be recognized from edges when viewed in the major, but not minor, groove
|
|
|
Do RNA pol and DNA pol needs primers?
|
RNA pol does NOT, DNA pol DOES.
|
|
|
Carboxy-terminal domain: Significance
|
The carboxy-terminal domain of RNA polymerase II. Other proteins often bind the C-terminal domain of RNA polymerase in order to activate polymerase activity. It is the protein domain which is involved in the initiation of DNA transcription (phosphorylation of this region permits escape from the promoter and activates the beginning of transcription), the capping of the RNA transcript, and attachment to the spliceosome for RNA splicing.
|